Citation: Chunyang Qin, Yuming Chen, Xia Wang. Global dynamics of a delayed diffusive virus infection model with cell-mediated immunity and cell-to-cell transmission[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4678-4705. doi: 10.3934/mbe.2020257

| [1] | M. A. Nowak, S. Bonhoeffer, A. M. Hill, R. Boehme, H. C. Thomas, H. McDade, Viral dynamics in hepatitis B virus infection, Proc. Natl. Acad. Sci. USA, 93 (1996), 4398-4402. |
| [2] | M. A. Nowak, R. M. May, Virus dynamics: Mathematical principles of immunology and virology, Oxford University Press, 2000. |
| [3] | J. Guedj, A. U. Neumann, Understanding hepatitis C viral dynamics with direct-acting antiviral agents due to the interplay between intracellular replication and cellular infection dynamics, J. Theoret. Biol., 267 (2010), 330-340. |
| [4] | L. Rong, A. S. Pereslon, Mathematical analysis of multiscale models for hepatitis C virus dynamics under therapy with direct-acting antiviral agents, Math. Biosci., 245 (2013), 22-30. |
| [5] | R. V. Culshaw, S. Ruan, G. Webb, A mathematical model of cell-to-cell spread of HIV-1 that includes a time delay, J. Math. Biol., 46 (2003), 425-444. |
| [6] | P. K. Srivastava, M. Banerjee, P. Chandra, A primary infection model for HIV and immune response with two discrete time delays, Differ. Equ. Dyn. Syst., 18 (2010), 385-399. |
| [7] | X. Wang, X. Song, S. Tang, L. Rong, Dynamics of an HIV model with multiple infection stages and treatment with different drug classes, Bull. Math. Biol., 78 (2016), 322-349. |
| [8] | X. Wang, G. Mink, D. Lin, X. Song, L. Rong, Influence of raltegravir intensification on viral load and 2-LTR dynamics in HIV patients on suppressive antiretroviral therapy, J. Theoret. Biol., 416 (2017), 16-27. |
| [9] | M. Y. Li, H. Shu, Impact of intracellular delays and target-cell dynamics on in vivo viral infections, SIAM J. Appl. Math., 70 (2010), 2434-2448. |
| [10] | M. Y. Li, H. Shu, Global dynamics of a mathematical model for HTLV-I infection of CD4+ T cells with delayed CTL response, Nonlinear Anal. Real World Appl., 13 (2012), 1080-1092. |
| [11] | H. Zhao, S. Liu, A mathematical model of HTLV-I infection with nonlinear incidence and two time delays, Commun. Math. Biol. Neurosci., 2016 (2016). |
| [12] | M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74-79. |
| [13] | K. Allali, S. Harroudi, D. F. M. Torres, Analysis and optimal control of an intracellular delayed HIV model with CTL immune response, Math. Comput. Sci., 12 (2018), 111-127. |
| [14] | X. Wang, Y. Tao, Lyapunov function and global properties of virus dynamics with CTL immune response, Int. J. Biomath., 1 (2008), 443-448. |
| [15] | J. Li, K. Men, Y. Yang, D. Li, Dynamical analysis on a chronic hepatitis C virus infection model with immune response, J. Theoret. Biol., 365 (2015), 337-346. |
| [16] | A. M. Elaiw, A. A. Raezah, K. Hattaf, Stability of HIV-1 infection with saturated virus-target and infected-target incidences and CTL immune response, Int. J. Biomath., 10 (2017), 1750070. |
| [17] | A. V. M. Herz, S. Bonhoeffer, R. M. Anderson, R. M. May, M. A. Nowak, Viral dynamics in vivo: limitations on estimates of intracellular delay and virus decay, Proc. Natl. Acad. Sci. USA, 93 (1996), 7247-7251. |
| [18] | H. Shu, L. Wang, J. Watmough, Global stability of a nonlinear viral infection model with infinitely distributed intracellular delays and CTL immune responses, SIAM J. Appl. Math., 73 (2013), 1280-1302. |
| [19] | Y. Liu, C. Wu, Global dynamics for an HIV infection model with Crowley-Martin functional response and two distributed delays, J. Syst. Sci. Complex., 31 (2018), 385-395. |
| [20] | H. Zhu, X. Zou, Dynamics of a HIV-1 infection model with cell-mediated immune response and intracellular delay, Discrete Contin. Dyn. Syst. Ser. B, 12 (2009), 511-524. |
| [21] | X. Wang, S. Liu, A class of delayed viral models with saturation infection rate and immune response, Math. Methods Appl. Sci., 36 (2013), 125-142. |
| [22] | B. Li, Y. Chen, X. Lu, S. Liu, A delayed HIV-1 model with virus waning term, Math. Biosci. Eng., 13 (2016), 135-157. |
| [23] | O. T. Fackler, T. T. Murooka, A. Imle, T. R. Mempel, Adding new dimensions: towards an integrative understanding of HIV-1 spread, Nat. Rev. Microbiol., 12 (2014), 563-574. |
| [24] | K. Wang, W. Wang, Propagation of HBV with spatial dependence, Math. Biosci., 210 (2007), 78-95. |
| [25] | R. Xu, Z. Ma, An HBV model with diffusion and time delay, J. Theoret. Biol., 257 (2009), 499-509. |
| [26] | C. C. McCluskey, Y. Yang, Global stability of a diffusive virus dynamics model with general incidence function and time delay, Nonlinear Anal. Real World Appl., 25 (2015), 64-78. |
| [27] | C. M. Brauner, D. Jolly, L. Lorenzi, R. Thiebaut, Heterogeneous viral environment in an HIV spatial model, Discrete Contin. Dyn. Syst. Ser. B, 15 (2011), 545-572. |
| [28] | Y. Zhang, Z. Xu, Dynamics of a diffusive HBV model with delayed Beddington-DeAngelis response, Nonlinear Anal. Real World Appl., 15 (2014), 118-139. |
| [29] | Y. Yang, Y. Xu, Global stability of a diffusive and delayed virus dynamics model with BeddingtonDeAngelis incidence function and CTL immune response, Comput. Math. Appl., 71 (2016), 922-930. |
| [30] | W. Hübner, G. P. McNerney, P. Chen, B. M. Dale1, R. E. Gordon, F. Y. S. Chuang, et al., Quantitative 3D video microscopy of HIV transfer across T cell virological synapses, Science, 323 (2009), 1743-1747. |
| [31] | P. Zhong, L. M. Agosto, J. B. Munro, W. Mothes, Cell-to-cell transmission of viruses, Curr. Opin. Virol., 3 (2013), 44-50. |
| [32] | N. Martin, Q. Sattentau, Cell-to-cell HIV-1 spread and its implications for immune evasion, Curr. Opin. HIV AIDS, 4 (2009), 143-149. |
| [33] | C. Zhang, S. Zhou, E. Groppelli, P. Pellegrino, I. Williams, P. Borrow, Hybrid spreading mechanisms and T cell activation shape the dynamics of HIV-1 infection, PLoS Comput. Biol., 11 (2015), e1004179. |
| [34] | A. Sigal, J. T. Kim, A. B. Balazs, E. Dekel, A. Mayo, R. Milo, et al., Cell-to-cell spread of HIV permits ongoing replication despite antiretroviral therapy, Nature, 477 (2011), 95-98. |
| [35] | X. Lai, X. Zou, Modeling HIV-1 virus dynamics with both virus-to-cell infection and cell-to-cell transmission, SIAM J. Appl. Math., 74 (2014), 898-917. |
| [36] | X. Lai, X. Zou, Modeling cell-to-cell spread of HIV-1 with logistic target cell growth, J. Math. Anal. Appl., 426 (2015), 563-584. |
| [37] | X. Wang, S. Tang, X. Song, L. Rong, Mathematical analysis of an HIV latent infection model including both virus-to-cell infection and cell-to-cell transmission, J. Biol. Dyn., 11 (2017), 455-483. |
| [38] | H. Shu, Y. Chen, L. Wang, Impacts of the cell-free and cell-to-cell infection modes on viral dynamics, J. Dyn. Differ. Equ., 30 (2018), 1817-1836. |
| [39] | A. Debadatta, B. Nandadulal, Analysis and computation of multi-pathways and multi-delays HIV-1 infection model, Appl. Math. Model., 54 (2018), 517-536. |
| [40] | X. Song, A. U. Neumann, Global stability and periodic solution of the viral dynamics, J. Math. Anal. Appl., 329 (2007), 281-297. |
| [41] | X. Wang, Y. Tao, X. Song, Global stability of a virus dynamics model with Beddington-DeAngelis incidence rate and CTL immune response, Nonlinear Dynam., 66 (2011), 825-830. |
| [42] | H. Sun, J. Wang, Dynamics of a diffusive virus model with general incidence function, cell-to-cell transmission and time delay, Comput. Math. Appl., 77 (2019), 284-301. |
| [43] | R. H. Martin, H. L. Smith, Abstract functional-differential equations and reaction-diffusion systems, Trans. Amer. Math. Soc., 321 (1990), 1-44. |
| [44] | Y. Gao, J. Wang, Threshold dynamics of a delayed nonlocal reaction-diffusion HIV infection model with both cell-free and cell-to-cell transmissions, J. Math. Anal. Appl., 488 (2020), 124047. |
| [45] | Y. Lou, X. Zhao, A reaction-diffusion malaria model with incubation period in the vector population, J. Math. Biol., 62 (2011), 543-568. |
| [46] | M. H. Protter, H. F. Weinberger, Maximum principles in differential equations, Springer-Verlag, 1984. |
| [47] | J. Wu, Theory and applications of partial functional differential equations, Springer, New York, 1996. |
| [48] | J. K. Hale, Asymptotic behavior of dissipative systems, American Mathematical Society, Providence, 1988. |
| [49] | W. Wang, X. Zhao, Basic reproduction numbers for reaction-diffusion epidemic models, SIAM J. Appl. Dyn. Syst., 11 (2012), 1652-1673. |
| [50] | J. K. Hale, S. M. Verduyn Lunel, Introduction to functional differential equations, Springer-Verlag, 1993. |
| [51] | J. LaSalle, S. Lefschetz, Stability by Lyapunov's direct method, with applications, in Mathematics in Science and Engineering, Academic Press, 1961. |
| [52] | H. R. Thieme, Convergence results and a Poincaré-Bendixson trichotomy for asymptotically autonomous differential equations, J. Math. Biol., 30 (1992), 755-763. |
| [53] | H. L. Smith, X. Zhao, Robust persistence for semidynamical systems, Nonlinear Anal., 47 (2001), 6169-6179. |
| [54] | P. W. Nelson, J. D. Murray, A. S. Perelson, A model of HIV-1 pathogenesis that includes an intracellular delay, Math. Biosci., 163 (2000), 201-215. |
| [55] | K. A. Pawelek, S. Liu, F. Pahlevani, L. Rong, A model of HIV-1 infection with two time delays: mathematical analysis and comparison with patient data, Math. Biosci., 235 (2012), 98-109. |
| [56] | H. McCallum, N. Barlow, J. Hone, How should pathogen transmission be modelled? Trends Ecol. Evol., 16 (2001), 295-300. |
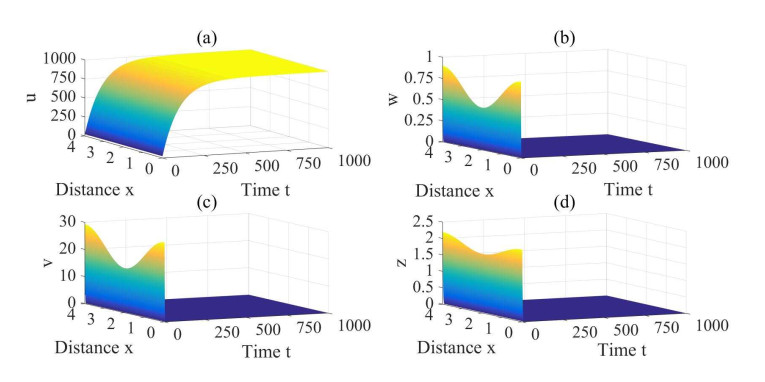
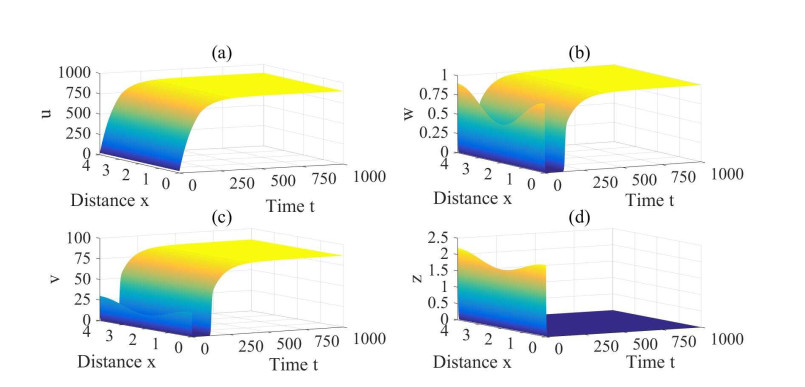
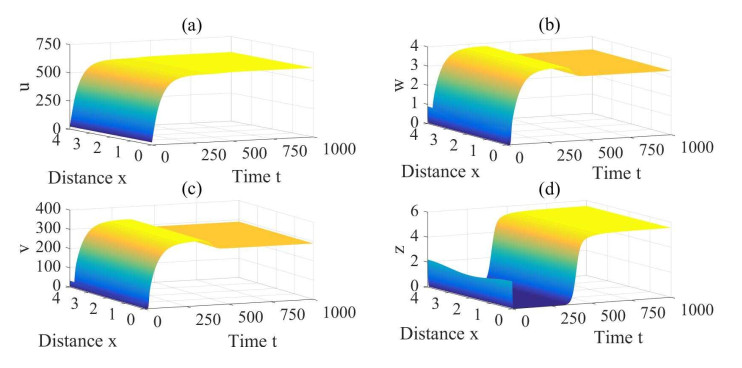
Figures(3) / Tables(1)

Chunyang Qin, Yuming Chen, Xia Wang. Global dynamics of a delayed diffusive virus infection model with cell-mediated immunity and cell-to-cell transmission[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4678-4705. doi: 10.3934/mbe.2020257







 DownLoad:
DownLoad: