Citation: Maoxing Liu, Shushu He, Yongzheng Sun. The impact of media converge on complex networks on disease transmission[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6335-6349. doi: 10.3934/mbe.2019316

| [1] | M. E. J. Newman, Spread of epidemic disease on networks, Phys. Rev. E, 66 (2002), 016128. |
| [2] | M. E. J. Newman, The structure and function of complex networks, SIAM Rev., 45 (2003), 167–256. |
| [3] | M. J. Keeling, The implications of network structure for epidemic dynamics, Theor. Popul. Biol., 67 (2005), 1–8. |
| [4] | R. Pastor-Satorras and A. Vespignani, Epidemic spreading in scale-free networks, Phys. Rev. Lett., 86 (2001), 3200–3203. |
| [5] | T. Zhou, J. G. Liu, W. J. Bai, et al., Behaviors of susceptible-infected epidemics on scale-free networks with identical infectivity, Phys. Rev. E, 74 (2006), 056109. |
| [6] | M. Liu and Y. Chen, An SIRS model with differential susceptibility and infectivity on uncorrelated networks, Math. Biosci. Eng., 12 (2015), 415–429. |
| [7] | J. Liu and T. Zhang, Epidemic spreading of an SEIRS model in scale-free networks, Commun. Nonlinear Sci. Numer. Simul., 16 (2011), 3375–3384. |
| [8] | R. Pastor-Satorras and A. Vespignani, Immunization of complex networks, Phys. Rev. E, 65 (2002), 036104. |
| [9] | A. d'Onofrio, A note on the global behaviour of the network-based SIS epidemic model, Nonlinear Anal., 9 (2008), 1567–1572. |
| [10] | J. Lou and T. Ruggeri, The dynamics of spreading and immune strategies of sexually transmitted diseases on scale-free network, J. Math. Anal. Appl., 365 (2010), 210–219. |
| [11] | W. J. Reed, A stochastic model for the spread of a sexually transmitted disease which results in a scale-free network, Math. Biosci., 201 (2006), 3–14. |
| [12] | M. Liu and J. Ruan, Modelling the spread of sexually transmitted diseases on scale-free networks, Chinese Physics B, 18 (2009), 2115–2120. |
| [13] | S. Funk, M. Salathé and V. A. A. Jansen, Modelling the influence of human behaviour on the spread of infectious diseases: a review, J. R. Soc. Interf., 7 (2010), 1247–1256. |
| [14] | R. Liu, J. Wu and H. Zhu, Media/psychological impact on multiple outbreaks of emerging infec-tious diseases, Comput. Math. Methods Med., 8 (2007), 153–164. |
| [15] | J. Tchuenche, N. Dube, C. Bhunu, et al., The impact of media coverage on the transmission dynamics of human influenza, BMC Public Health, 11 (2011), S5. |
| [16] | M. Liu, E. Liz and G. Röst, Endemic bubbles generated by delayed behavioral response–global stability and bifurcation switches in an SIS model, SIAM J. Appl. Math., 75 (2015), 75–91. |
| [17] | L. Zuo, M. Liu and J. Wang, The impact of awareness programs with recruitment and delay on the spread of an epidemic, Math. Prob. Eng., 2014 (2014), 235935. |
| [18] | S. Collinson and J. M. Heffernan, Modelling the effects of media during an influenza epidemic, BMC Public Health, 14 (2014), 1–10. |
| [19] | K. A. Pawelek, A. Oeldorf-Hirsch and L. Rong, Modeling the impact of twitter on influenza epidemics, Math. Biosci. Eng., 11 (2014), 1337–1356. |
| [20] | D. Greenhalgh, S. Rana, S. Samanta, et al., Awareness programs control infectious disease-Multiple delay induced mathematical model, Appl. Math. Comput., 251 (2015), 539–563. |
| [21] | Y. Wang, J. Cao, Z. Jin, et al., Impact of media coverage on epidemic spreading in complex networks, Phys. A, 392 (2013), 5824–5835. |
| [22] | X. Yuan, Y. Xue and M. Liu, Analysis of an epidemic model with awareness programs by media on complex networks, Chaos Soli. Frac., 48 (2013), 1–11. |
| [23] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equi-libria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [24] | P. De Leenheer and H. L. Smith, Virus dynamics: A global analysis, SIAM J. Appl. Math., 63 (2003), 1313–1327. |
| [25] | H. R. Thieme, Persistence under relaxed point-dissipativity (with application to an endemic model), SIAM J. Math. Anal., 24 (1993), 407–435. |
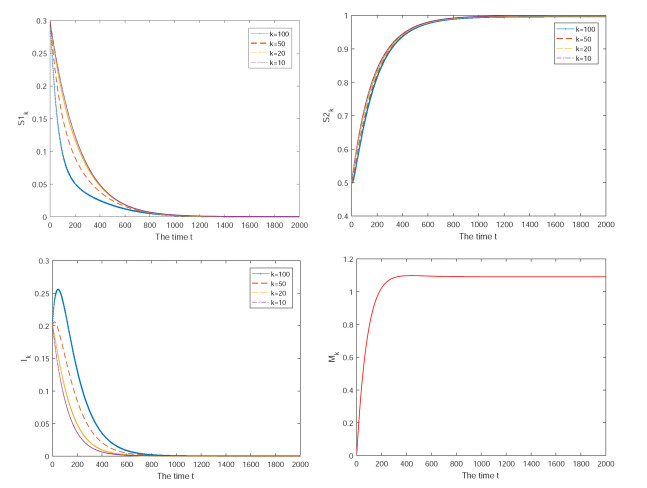
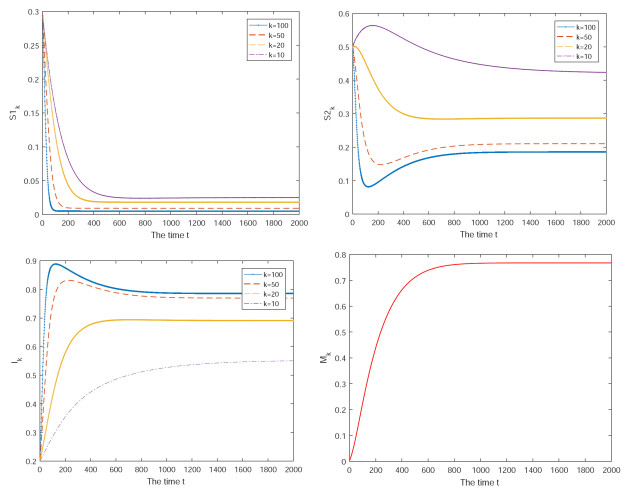
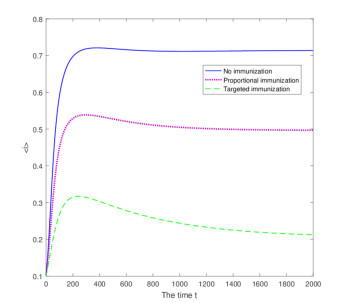
Figures(3)

Maoxing Liu, Shushu He, Yongzheng Sun. The impact of media converge on complex networks on disease transmission[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6335-6349. doi: 10.3934/mbe.2019316







 DownLoad:
DownLoad: