Citation: Eduardo González-Olivares, Claudio Arancibia-Ibarra, Alejandro Rojas-Palma, Betsabé González-Yañez. Bifurcations and multistability on the May-Holling-Tanner predation model considering alternative food for the predators[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4274-4298. doi: 10.3934/mbe.2019213

| [1] | S. Yu, Global asymptotic stability of a predator-prey model with modified Leslie-Gower and Holling-type II schemes, Discrete Dyn. Nat. Soc., 2012, 208167. |
| [2] | Z. Zhao, L. Yang and L. Chen, Impulsive perturbations of a predator-prey system with modified Leslie-Gower and Holling type II schemes, J. App. Math. Comput., 35 (2011), 119–134. |
| [3] | D. K. Arrowsmith and C. M. Place, Dynamical Systems. Differential equations, maps and chaotic behaviour, Chapman and Hall, 1992. |
| [4] | E. González-Olivares, L. M. Gallego-Berrío, B. González-Yañez, et al., Consequences of weak Allee effect on prey in the May-Holling-Tanner predator-prey model, Math. Meth. Appl. Sci., 38 (2015), 5183–5196. |
| [5] | E. Sáez and E. González-Olivares, Dynamics on a predator-prey model, SIAM J. Appl. Math., 59 (1999), 1867–1878. |
| [6] | J. T. Tanner, The stability and the intrinsic growth rate of prey and predator population, Ecology, 56 (1975), 855–867. |
| [7] | P. H. Leslie and J. C. Gower, The properties of a stochastic model for the predator-prey type of interaction between two species, Biometrika, 47 (1960), 219–234. |
| [8] | P. H. Leslie, Some further notes on the use of matrices in Population Mathematics, Biometrika, 35 (1948), 213–245. |
| [9] | R. M. May, Stability and complexity in model ecosystems (2nd edition), Princeton University Press, 2001. |
| [10] | P. Turchin, Complex population dynamics. A theoretical/empirical synthesis, Monographs in Population Biology 35, Princeton University Press (2003). |
| [11] | J. Mena-Lorca, E. González-Olivares and B. González-Yañez, The Leslie-Gower predator-prey model with Allee effect on prey: A simple model with a rich and interesting dynamics, in: R. Mondaini (Ed.), Proceedings of the 2006 International Symposium on Mathematical and Computational Biology BIOMAT 2006, E-papers Servic ¸os Editoriais Ltda., Rio de Janeiro, 2007, 105–132. |
| [12] | P. Aguirre, E. González-Olivares and E. Sáez, Two limit cycles in a Leslie-Gower predator-prey model with additive Allee effect, Nonl Anal. Real World Appl., 10 (2009), 1401–1416. |
| [13] | M. A. Aziz-Alaoui and M. Daher Okiye, Boundedness and global stability for a predator-prey model with modified Leslie-Gower and Holling-type II schemes, Appl. Math. Lett., 16 (2003), 1069–1075. |
| [14] | J. B. Collings, The effect of the functional response on the bifurcation behavior of a mite predator- prey interaction model, J. Math. Biol., 36 (1997), 149–168. |
| [15] | K. Q. Lan and C. R. Zhu, Phase portraits, Hopf bifurcations and limit cycles of the Holling-Tanner models for predator-prey interactions, Nonlinear Anal. Real World Appl., 12 (2011), 1961–1973. |
| [16] | J. D. Murray, Mathematical Biology: I. An Introduction (3ird Edition) Springer, 2001. |
| [17] | D. J. Wollkind, J. B.Collings and J. A. Logan, Metastability in a temperature-dependent model system for predator-prey mite outbreak interactions on fruit trees, Bull. Math. Biol., 50 (1988), 379–409. |
| [18] | S.-B. Hsu and T.-W. Huang, Global stability for a class of predator-prey systems, SIAM J. Appl. Math., 55 (1995), 763–783. |
| [19] | P. Aguirre, E. González-Olivares and E. Sáez, Three limit cycles in a Leslie-Gower predator-prey model with additive Allee effect, SIAM J. Appl. Math., 69 (2009), 1244–1269. |
| [20] | B. González-Yañez, E. González-Olivares and J. Mena-Lorca, Multistability on a Leslie-Gower Type predator-prey model with nonmonotonic functional response, In R. Mondaini and R. Dilao (eds.), BIOMAT 2006 - International Symposium on Mathematical and Computational Biology, World Scientific Co. Pte. Ltd., (2007), 359–384. |
| [21] | I. Hanski, H. Hentonnen, E. Korpimaki, et al., Small-rodent dynamics and predation, Ecology, 82 (2001), 1505–1520. |
| [22] | C. Arancibia-Ibarra and E. González-Olivares, A modified Leslie-Gower predator-prey model with hyperbolic functional response and Allee effect on prey, In R. Mondaini (Ed.) BIOMAT 2010 International Symposium on Mathematical and Computational Biology, World Scientific Co. Pte. Ltd., Singapore, (2011), 146–162. |
| [23] | P. Feng and Y. Kang, Dynamics of a modified Leslie-Gower model with double Allee effects, Nonlinear Dyn., 80 (2015), 1051–1062. |
| [24] | A. Singh and S. Gakkhar, Stabilization of modified Leslie-Gower prey-predator model, Differ. Equ. Dyn. Syst., (2014), 239–249. |
| [25] | A. Korobeinikov, A Lyapunov function for Leslie-Gower predator-prey models, Appl. Math. Lett., 14 (2001), 697–699. |
| [26] | A. D. Bazykin, Nonlinear Dynamics of interacting populations, World Scientific Publishing Co. Pte. Ltd., 1998. |
| [27] | R. J. Taylor, Predation, Chapman and Hall, 1984. |
| [28] | W. M. Getz, A hypothesis regarding the abruptness of density dependence and the growth rate populations, Ecology, 77 (1996), 2014–2026. |
| [29] | R. Yuan, W. Jiang and Y. Wang, Saddle-node-Hopf bifurcation in a modified Leslie-Gower predator-prey model with time-delay and prey harvesting, J. Math. Anal. Appl., 422 (2015), 1072– 1090. |
| [30] | Q. Yue, Dynamics of a modified Leslie-Gower predator-prey model with Holling-type II schemes and a prey refuge, Springer Plus, 5 (2016), 461. |
| [31] | E. González-Olivares, P. Tintinago-Ruiz and A. Rojas-Palma, A Leslie-Gower type predator-prey model with sigmoid funcional response, Internat. J. Comp. Math., 93 (2015), 1895–1909. |
| [32] | C. Chicone, Ordinary differential equations with applications (2nd edition), Texts in Applied Mathematics 34, Springer 2006. |
| [33] | F. Dumortier, J. Llibre and J. C. Artés, Qualitative theory of planar differential systems, Springer 2006. |
| [34] | H. I. Freedman, Deterministic Mathematical Model in Population Ecology, Marcel Dekker, 1980. |
| [35] | L. Nie, Z. Teng, L. Hua and J. Peng, Qualitative analysis of a modified Leslie-Gower and Holling- type II predator prey model with state dependent impulsive effects, Nonlinear Anal. Real World Appl., 11 (2010), 1364–1373. |
| [36] | Y. Tian and P. Weng, Stability analysis of diffusive predator-prey model with modified Leslie- Gower and Holling-type II schemes, Acta Appl. Math., 114 (2011), 173–192. |
| [37] | E. González-Olivares, J. Mena-Lorca, A. Rojas-Palma, et al., Dynamical complexities in the Leslie-Gower predator-prey model as consequences of the Allee effect on prey, Appl. Math. Model., 35 (2011), 366–381. |
| [38] | C. Arancibia-Ibarra and E. González-Olivares, The Holling-Tanner model considering an alternative food for predator, In J. Vigo-Aguiar (Ed.) Proceedings of the 2015 International Conference on Computational and Mathematical Methods in Science and Engineering CMMSE 2015, 1 (2015), 130–141. |
| [39] | E. González-Olivares, C. Arancibia-Ibarra, B. González-Yañez, et al., Dynamics of a Leslie- Gower predation model considering a generalist predator and the hyperbolic functional response, submitted (2019). |
| [40] | A. Dhooge, W. Govaerts and Y. Kuznetsov, Matcont: a matlab package for numerical bifurcation analysis of ODES, ACM Trans. Math. Soft. (TOMS), 29 (2003), 141–164. |
| [41] | V. A. Gaiko, Global Bifurcation Theory and Hilbert's Sixteenth Problem, in: Mathematics and its Applications, vol. 559, Kluwer Academic Publishers, 2003. |
| [42] | B-S. Goh, Management and analysis of biological populations, Elsevier Scientific Publishing Company, 1980. |
| [43] | A. Rantzer, A dual to Lyapunov's stability theorem, Syst. Control Lett., 42 (2001), 161–168. |
| [44] | Y. A. Kuznetsov, Elements of applied bifurcation theory (3rd ed) Springer-Verlag, 2004. |
| [45] | P. Monzón, Almost global attraction in planar systems, Syst. Control Lett, 54 (2005), 753–758. |
| [46] | S. Wolfram, Mathematica: A System for Doing Mathematics by Computer (2nd edition), Wolfram Research, Addison Wesley 1991. |
| [47] | D. Xiao and S. Ruan, Bogdanov-Takens bifurcations in predator-prey systems with constant rate harvesting, Field Inst. Comm., 21 (1999), 493–506. |
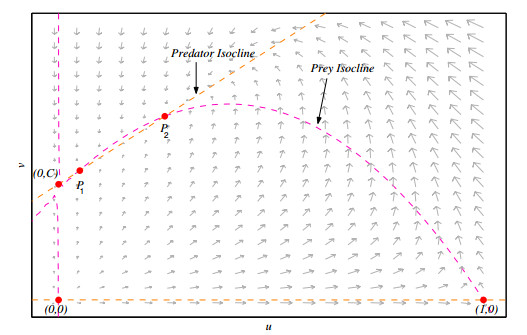
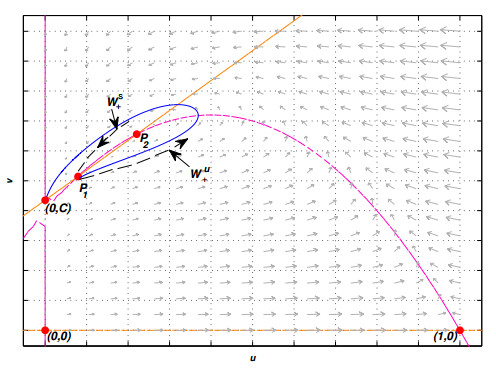
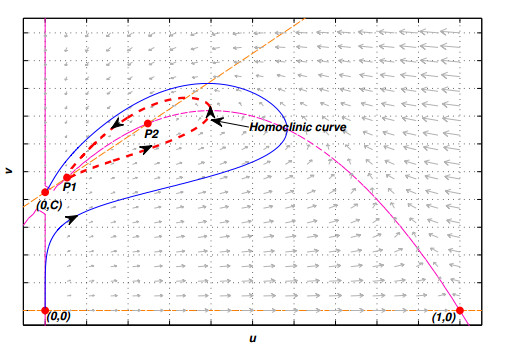
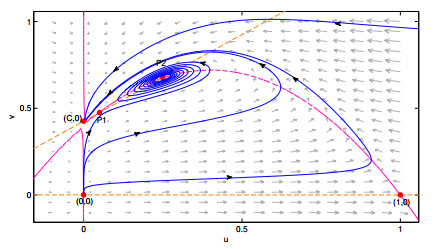
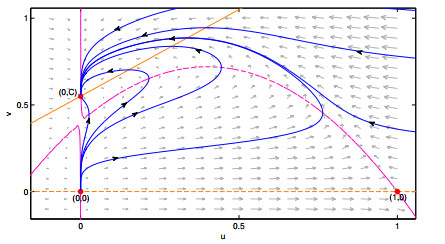
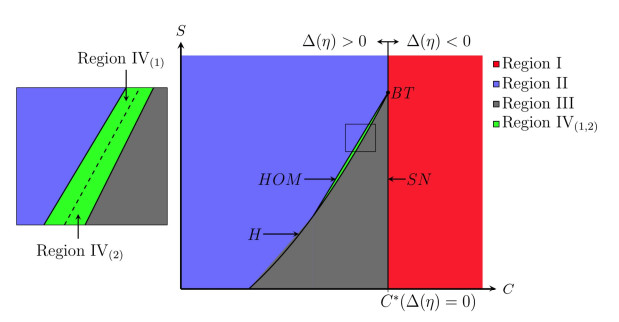
Figures(10) / Tables(2)

Eduardo González-Olivares, Claudio Arancibia-Ibarra, Alejandro Rojas-Palma, Betsabé González-Yañez. Bifurcations and multistability on the May-Holling-Tanner predation model considering alternative food for the predators[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4274-4298. doi: 10.3934/mbe.2019213







 DownLoad:
DownLoad: