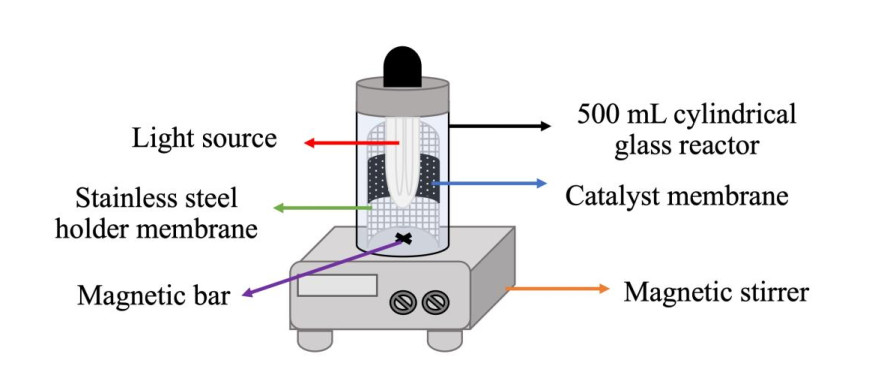
In this study, Mn-doped ZnO composite carbon nanofibers (Mn-ZnO/CNFs) were prepared via a simple blending and electrospinning (ES) method, followed by a thermal treatment. These fibers were used to investigate the photocatalytic degradation of an organic dye under UV and visible light irradiation. The results showed that Mn-ZnO/CNFs were successfully prepared under the same conditions used for CNFs preparation conditions, which induced a morphological change from a smooth to a rough surface compared to the CNFs. Energy dispersive X-ray (EDX), x-ray diffraction (XRD), and x-ray photoelectron spectroscopy (XPS) analyses confirmed the formation of Mn-doped ZnO on the CNFs' surface. Furthermore, the addition of the catalyst significantly increased in the specific surface area, and a N2 adsorption-desorption isotherm analysis revealed that all samples had mesoporous characteristics with a type IV isotherm index. The photocatalytic activity of the Mn-ZnO/CNFs carbonized at 650 ℃ using methylene blue (MB) dye as a model pollutant was investigated. All prepared samples effectively removed the MB with a degradation rate of 70-90%. The kinetic reaction rate was described using the simplified Langmuir-Hinshelwood equation. Overall, the CNFs and composites nanofibers developed through moderate thermal treatment processes possessed a high specific surface area and oxygen vacancy, enabling their potential use as adsorbents and as a catalyst support for reactions at room-to-elevated temperatures, as well as photocatalysts for the removal of organic contaminants.
Citation: Krittiya Singcharoen, Ratthapol Rangkupan, Soontree Khuntong, Thanakorn Wasanapiarnpong. Enhancing photocatalytic efficiency with Mn-doped ZnO composite carbon nanofibers for organic dye degradation[J]. AIMS Environmental Science, 2024, 11(2): 324-341. doi: 10.3934/environsci.2024015
In this study, Mn-doped ZnO composite carbon nanofibers (Mn-ZnO/CNFs) were prepared via a simple blending and electrospinning (ES) method, followed by a thermal treatment. These fibers were used to investigate the photocatalytic degradation of an organic dye under UV and visible light irradiation. The results showed that Mn-ZnO/CNFs were successfully prepared under the same conditions used for CNFs preparation conditions, which induced a morphological change from a smooth to a rough surface compared to the CNFs. Energy dispersive X-ray (EDX), x-ray diffraction (XRD), and x-ray photoelectron spectroscopy (XPS) analyses confirmed the formation of Mn-doped ZnO on the CNFs' surface. Furthermore, the addition of the catalyst significantly increased in the specific surface area, and a N2 adsorption-desorption isotherm analysis revealed that all samples had mesoporous characteristics with a type IV isotherm index. The photocatalytic activity of the Mn-ZnO/CNFs carbonized at 650 ℃ using methylene blue (MB) dye as a model pollutant was investigated. All prepared samples effectively removed the MB with a degradation rate of 70-90%. The kinetic reaction rate was described using the simplified Langmuir-Hinshelwood equation. Overall, the CNFs and composites nanofibers developed through moderate thermal treatment processes possessed a high specific surface area and oxygen vacancy, enabling their potential use as adsorbents and as a catalyst support for reactions at room-to-elevated temperatures, as well as photocatalysts for the removal of organic contaminants.

| [1] |
Al-Tohamy R, Ali SS, Li F, et al. (2022) A critical review on the treatment of dye-containing wastewater: Ecotoxicological and health concerns of textile dyes and possible remediation approaches for environmental safety. Ecotoxicol Environ Saf 231: 113160. https://doi.org/10.1016/j.ecoenv.2021.113160 doi: 10.1016/j.ecoenv.2021.113160

|
| [2] |
Piaskowski K, Świderska-Dąbrowska R, Zarzycki PK (2018) Dye Removal from Water and Wastewater Using Various Physical, Chemical, and Biological Processes. J AOAC Int 101: 1371–1384. https://doi.org/10.5740/jaoacint.18-0051 doi: 10.5740/jaoacint.18-0051
|
| [3] |
Peramune D, Manatunga DC, Dassanayake RS, et al. (2022) Recent advances in biopolymer-based advanced oxidation processes for dye removal applications: A review. Environ Res 215: 114242. https://doi.org/10.1016/j.envres.2022.114242 doi: 10.1016/j.envres.2022.114242
|
| [4] |
Buzzetti L, Crisenza GEM, Melchiorre P (2019) Mechanistic Studies in Photocatalysis. Angewandte Chemie Int Ed 58: 3730–3747. https://doi.org/10.1002/anie.201809984 doi: 10.1002/anie.201809984
|
| [5] |
Ângelo J, Andrade L, Madeira LM, et al. (2013) An overview of photocatalysis phenomena applied to NOx abatement. J Environ Manage 129: 522–539. https://doi.org/10.1016/j.jenvman.2013.08.006 doi: 10.1016/j.jenvman.2013.08.006
|
| [6] |
Sarkodie B, Amesimeku J, Frimpong C, et al. (2023) Photocatalytic degradation of dyes by novel electrospun nanofibers: A review. Chemosphere 313: 137654. https://doi.org/10.1016/j.chemosphere.2022.137654 doi: 10.1016/j.chemosphere.2022.137654
|
| [7] |
Pomilla FR, García-López EI, Marcì G, et al. (2021) Heterogeneous photocatalytic materials for sustainable formation of high-value chemicals in green solvents. Mater Today Sustain 13: 100071. https://doi.org/10.1016/j.mtsust.2021.100071 doi: 10.1016/j.mtsust.2021.100071
|
| [8] |
Jadoun S, Yáñez J, Mansilla HD, et al. (2022) Conducting polymers/zinc oxide-based photocatalysts for environmental remediation: a review. Environ Chem Lett 20: 2063–2083. https://doi.org/10.1007/s10311-022-01398-w doi: 10.1007/s10311-022-01398-w
|
| [9] |
Lee KM, Lai CW, Ngai KS, et al. (2016) Recent developments of zinc oxide based photocatalyst in water treatment technology: A review. Water Res 88: 428–448. https://doi.org/10.1016/j.watres.2015.09.045 doi: 10.1016/j.watres.2015.09.045
|
| [10] | Mohamed KM, Benitto JJ, Vijaya JJ, et al. (2023) Recent Advances in ZnO-Based Nanostructures for the Photocatalytic Degradation of Hazardous, Non-Biodegradable Medicines. Crystals 13. https://doi.org/10.3390/cryst13020329 |
| [11] | Mao T, Liu M, Lin L, et al. (2022) A Study on Doping and Compound of Zinc Oxide Photocatalysts. Polymers 14. https://doi.org/10.3390/polym14214484 |
| [12] |
Ab Wahab N, Sairi NA, Alias Y (2022) Photocatalytic activities enhancement of manganese doped ZnO by decoration on CNT for degradation of organic pollutants under solar irradiation. Appl Phys A Mater Sci Process 128: 1–10. https://doi.org/10.1007/s00339-021-05160-x doi: 10.1007/s00339-021-05160-x
|
| [13] |
Thakur P, Chahar D, Taneja S, et al. (2020) A review on MnZn ferrites: Synthesis, characterization and applications. Ceram Int 46: 15740–15763. https://doi.org/10.1016/j.ceramint.2020.03.287 doi: 10.1016/j.ceramint.2020.03.287
|
| [14] |
Ullah R, Dutta J (2008) Photocatalytic degradation of organic dyes with manganese-doped ZnO nanoparticles. J Hazard Mater 156: 194–200. https://doi.org/10.1016/j.jhazmat.2007.12.033 doi: 10.1016/j.jhazmat.2007.12.033
|
| [15] |
Ayoub I, Kumar V, Abolhassani R, et al. (2022) Advances in ZnO: Manipulation of defects for enhancing their technological potentials. Nanotechnol Rev 11: 575–619. https://doi.org/10.1515/ntrev-2022-0035 doi: 10.1515/ntrev-2022-0035
|
| [16] |
Rauf MA, Meetani MA, Hisaindee S (2011) An overview on the photocatalytic degradation of azo dyes in the presence of TiO2 doped with selective transition metals. Desalination 276: 13–27. https://doi.org/10.1016/j.desal.2011.03.071 doi: 10.1016/j.desal.2011.03.071
|
| [17] |
Xu J-J, Lu Y-N, Tao F-F, et al. (2023) ZnO Nanoparticles Modified by Carbon Quantum Dots for the Photocatalytic Removal of Synthetic Pigment Pollutants. ACS Omega 8: 7845–7857. https://doi.org/10.1021/acsomega.2c07591 doi: 10.1021/acsomega.2c07591
|
| [18] |
Chabalala MB, Al-Abri MZ, Mamba BB, et al. (2021) Mechanistic aspects for the enhanced adsorption of bromophenol blue and atrazine over cyclodextrin modified polyacrylonitrile nanofiber membranes. Chem Eng Res Des 169: 19–32. https://doi.org/10.1016/j.cherd.2021.02.010 doi: 10.1016/j.cherd.2021.02.010
|
| [19] | Ferrone E, Araneo R, Notargiacomo A, et al. (2019) ZnO Nanostructures and Electrospun ZnO–Polymeric Hybrid Nanomaterials in Biomedical, Health, and Sustainability Applications. Nanomaterials 9. https://doi.org/10.3390/nano9101449 |
| [20] |
Asgari S, Mohammadi Ziarani G, Badiei A, et al. (2022) Electrospun composite nanofibers as novel high-performance and visible-light photocatalysts for removal of environmental pollutants: A review. Environ Res 215: 114296. https://doi.org/10.1016/j.envres.2022.114296 doi: 10.1016/j.envres.2022.114296
|
| [21] |
Li C, Yang J, Zhang L, et al. (2021) Carbon-based membrane materials and applications in water and wastewater treatment: a review. Environ Chem Lett 19: 1457–1475. https://doi.org/10.1007/s10311-020-01112-8 doi: 10.1007/s10311-020-01112-8
|
| [22] | Wang W, Chen Y, Wang N, et al. (2022) Multifunction ZnO/carbon hybrid nanofiber mats for organic dyes treatment via photocatalysis with enhanced solar-driven evaporation. Front Mater Sci 16. https://doi.org/10.1007/s11706-022-0623-7 |
| [23] | Wang Y, Hao X, Wang Z, et al. (2020) Facile fabrication of Mn2+- doped ZnO photocatalysts by electrospinning. R Soc Open Sci 7. https://doi.org/10.1098/rsos.191050 |
| [24] |
Baylan E, Altintas Yildirim O (2019) Highly efficient photocatalytic activity of stable manganese-doped zinc oxide (Mn:ZnO) nanofibers via electrospinning method. Mater Sci Semicond Process 103: 104621. https://doi.org/10.1016/j.mssp.2019.104621 doi: 10.1016/j.mssp.2019.104621
|
| [25] | Wang Y, Cheng J, Yu S, et al. (2016) Synergistic effect of N-decorated and Mn2+ doped ZnO nanofibers with enhanced photocatalytic activity. Sci Rep 6. https://doi.org/10.1038/srep32711 |
| [26] |
Samadi M, Zirak M, Naseri A, et al. (2016) Recent progress on doped ZnO nanostructures for visible-light photocatalysis. Thin Solid Films 605: 2–19. https://doi.org/10.1016/j.tsf.2015.12.064 doi: 10.1016/j.tsf.2015.12.064
|
| [27] |
Dai Z, Zhang CY, Guowen S, et al. (2020) Highly enhanced electrochemical cycling stabilities of hierarchical partially-embedded MnO/carbon nanofiber composites as supercapacitor electrodes. Mat Sci Eng B 262: 114684. https://doi.org/10.1016/j.mseb.2020.114684 doi: 10.1016/j.mseb.2020.114684
|
| [28] |
Suganthi K, Vinoth E, Sudha L, et al. (2023) Manganese (Mn2+) doped hexagonal prismatic zinc oxide (ZnO) nanostructures for chemiresistive NO2 sensor. Sens Actuators B Chem 380: 133293. https://doi.org/10.1016/j.snb.2023.133293 doi: 10.1016/j.snb.2023.133293
|
| [29] | Akram R, Fatima A, Almohaimeed ZM, et al. (2023) Photocatalytic Degradation of Methyl Green Dye Mediated by Pure and Mn-Doped Zinc Oxide Nanoparticles under Solar Light Irradiation. Adsorpt Sci Technol 2023. https://doi.org/10.1155/2023/5069872 |
| [30] |
Putri NA, Fauzia V, Iwan S, et al. (2018) Mn-doping-induced photocatalytic activity enhancement of ZnO nanorods prepared on glass substrates. Appl Surf Sci 439: 285–297. https://doi.org/10.1016/j.apsusc.2017.12.246 doi: 10.1016/j.apsusc.2017.12.246
|
| [31] |
Samuel E, Joshi B, Kim M-W, et al. (2019) Hierarchical zeolitic imidazolate framework-derived manganese-doped zinc oxide decorated carbon nanofiber electrodes for high performance flexible supercapacitors. Chem Eng J 371: 657–665. https://doi.org/10.1016/j.cej.2019.04.065 doi: 10.1016/j.cej.2019.04.065
|
| [32] |
Kim CH, Kim B-H (2015) Zinc oxide/activated carbon nanofiber composites for high-performance supercapacitor electrodes. J Power Sources 274: 512–520. https://doi.org/10.1016/j.jpowsour.2014.10.126 doi: 10.1016/j.jpowsour.2014.10.126
|
| [33] |
Luo Q, Yang X, Zhao X, et al. (2017) Facile preparation of well-dispersed ZnO/cyclized polyacrylonitrile nanocomposites with highly enhanced visible-light photocatalytic activity. Appl Catal B 204: 304–315. https://doi.org/10.1016/j.apcatb.2016.11.037 doi: 10.1016/j.apcatb.2016.11.037
|
| [34] |
Samadi M, Shivaee HA, Zanetti M, et al. (2012) Visible light photocatalytic activity of novel MWCNT-doped ZnO electrospun nanofibers. J Mol Catal A Chem 359: 42–48. https://doi.org/10.1016/j.molcata.2012.03.019 doi: 10.1016/j.molcata.2012.03.019
|
| [35] |
Samadi M, Shivaee HA, Pourjavadi A, et al. (2013) Synergism of oxygen vacancy and carbonaceous species on enhanced photocatalytic activity of electrospun ZnO-carbon nanofibers: Charge carrier scavengers mechanism. Appl Catal A Gen 466: 153–160. https://doi.org/10.1016/j.apcata.2013.06.024 doi: 10.1016/j.apcata.2013.06.024
|
| [36] |
Ning H, Xie H, Zhao Q, et al. (2017) Electrospinning ZnO/carbon nanofiber as binder-free and self-supported anode for Li-ion batteries. J Alloys Compd 722: 716–720. https://doi.org/10.1016/j.jallcom.2017.06.099 doi: 10.1016/j.jallcom.2017.06.099
|
| [37] |
Achouri F, Corbel S, Balan L, et al. (2016) Porous Mn-doped ZnO nanoparticles for enhanced solar and visible light photocatalysis. Mater Des 101: 309–316. https://doi.org/10.1016/j.matdes.2016.04.015 doi: 10.1016/j.matdes.2016.04.015
|
| [38] |
Liu L, Liu Z, Yang Y, et al. (2018) Photocatalytic properties of Fe-doped ZnO electrospun nanofibers. Ceram Int 44: 19998–20005. https://doi.org/10.1016/j.ceramint.2018.07.268 doi: 10.1016/j.ceramint.2018.07.268
|
| [39] | Alkaykh S, Mbarek A, Ali-Shattle EE (2020) Photocatalytic degradation of methylene blue dye in aqueous solution by MnTiO3 nanoparticles under sunlight irradiation. Heliyon 6: e03663. https://doi.org/10.1016/j.heliyon.2020.e03663 |
| [40] |
Wang D, Zhang M, Zhuang H, et al. (2017) The photocatalytic properties of hollow (GaN)1-x(ZnO)x composite nanofibers synthesized by electrospinning. Appl Surf Sci 396: 888–896. https://doi.org/10.1016/j.apsusc.2016.11.053 doi: 10.1016/j.apsusc.2016.11.053
|
| [41] |
Xu Z, Li X, Wang W, et al. (2016) Microstructure and photocatalytic activity of electrospun carbon nanofibers decorated by TiO2 nanoparticles from hydrothermal reaction/blended spinning. Ceram Int 42: 15012–15022. https://doi.org/10.1016/j.ceramint.2016.06.150 doi: 10.1016/j.ceramint.2016.06.150
|
| [42] |
Shah AP, Jain S, Mokale VJ, et al. (2019) High performance visible light photocatalysis of electrospun PAN/ZnO hybrid nanofibers. J Ind Eng Chem 77: 154–163. https://doi.org/10.1016/j.jiec.2019.04.03 doi: 10.1016/j.jiec.2019.04.03
|
| [43] |
Lakshmi K, Kadirvelu K, Mohan PS (2018) Photo-decontamination of p-nitrophenol using reusable lanthanum doped ZnO electrospun nanofiber catalyst. J Mater Sci Mater El 29: 12109–12117. https://doi.org/10.1007/s10854-018-9317-4 doi: 10.1007/s10854-018-9317-4
|








Figures(9) / Tables(4)

Krittiya Singcharoen, Ratthapol Rangkupan, Soontree Khuntong, Thanakorn Wasanapiarnpong. Enhancing photocatalytic efficiency with Mn-doped ZnO composite carbon nanofibers for organic dye degradation[J]. AIMS Environmental Science, 2024, 11(2): 324-341. doi: 10.3934/environsci.2024015







 DownLoad:
DownLoad: