Citation: Nick K. Olrichs, J. Bernd Helms. Novel insights into the function of the conserved domain of the CAP superfamily of proteins[J]. AIMS Biophysics, 2016, 3(2): 232-246. doi: 10.3934/biophy.2016.2.232

| [1] | Gibbs GM, Roelants K, O'Bryan MK (2008) The CAP superfamily: cysteine-rich secretory proteins, antigen 5, and pathogenesis-related 1 proteins--roles in reproduction, cancer, and immune defense. Endocr Rev 29: 865–897. |
| [2] | Gibbs GM, O'Bryan MK (2007) Cysteine rich secretory proteins in reproduction and venom. Soc Reprod Fertil Suppl 65: 261–267. |
| [3] | Yamazaki Y, Morita T (2004) Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 44: 227–231. |
| [4] | Choudhary V, Schneiter R (2012) Pathogen-Related Yeast (PRY) proteins and members of the CAP superfamily are secreted sterol-binding proteins. Proc Natl Acad Sci USA 109: 16882–16887. |
| [5] | Xu X, Francischetti IM, Lai R, et al. (2012) Structure of protein having inhibitory disintegrin and leukotriene scavenging functions contained in single domain. J Biol Chem 287: 10967–10976. |
| [6] | Kelleher A, Darwiche R, Rezende WC, et al. (2014) Schistosoma mansoni venom allergen-like protein 4 (SmVAL4) is a novel lipid-binding SCP/TAPS protein that lacks the prototypical CAP motifs. Acta Crystallogr D Biol Crystallogr 70: 2186–2196. |
| [7] | Milne TJ, Abbenante G, Tyndall JD, et al. (2003) Isolation and characterization of a cone snail protease with homology to CRISP proteins of the pathogenesis-related protein superfamily. J Biol Chem 278: 31105–31110. |
| [8] | Eberle HB, Serrano RL, Fullekrug J, et al. (2002) Identification and characterization of a novel human plant pathogenesis-related protein that localizes to lipid-enriched microdomains in the Golgi complex. J Cell Sci 115: 827–838. |
| [9] | Serrano RL, Kuhn A, Hendricks A, et al. (2004) Structural analysis of the human Golgi-associated plant pathogenesis related protein GAPR-1 implicates dimerization as a regulatory mechanism. J Mol Biol 339: 173–183. |
| [10] | Shoji-Kawata S, Sumpter R, Leveno M, et al. (2013) Identification of a candidate therapeutic autophagy-inducing peptide. Nature 494: 201–206. |
| [11] | Kuballa P, Nolte WM, Castoreno AB, et al. (2012) Autophagy and the immune system. Annu Rev Immunol 30: 611–646. |
| [12] | Eberle HB, Serrano RL, Fullekrug J, et al. (2002) Identification and characterization of a novel human plant pathogenesis-related protein that localizes to lipid-enriched microdomains in the Golgi complex. J Cell Sci 115: 827–838. |
| [13] | Zhou Q, Hao L, Huang W, et al. (2016) The Golgi-Associated Plant Pathogenesis-Related Protein GAPR-1 Enhances Type I Interferon Signaling Pathway in Response to Toll-Like Receptor 4. Inflammation 39: 706–717. |
| [14] | Baxter RM, Crowell TP, George JA, et al. (2007) The plant pathogenesis related protein GLIPR-2 is highly expressed in fibrotic kidney and promotes epithelial to mesenchymal transition in vitro. Matrix Biol 26: 20–29. |
| [15] | Aalberts M, van Dissel-Emiliani FM, van Adrichem NP, et al. (2012) Identification of distinct populations of prostasomes that differentially express prostate stem cell antigen, annexin A1, and GLIPR2 in humans. Biol Reprod 86: 82:1–8. |
| [16] | Van Galen J, Van Balkom BW, Serrano RL, et al. (2010) Binding of GAPR-1 to negatively charged phospholipid membranes: unusual binding characteristics to phosphatidylinositol. Mol Membr Biol 27: 81–91. |
| [17] | van Galen J, Olrichs NK, Schouten A, et al. (2012) Interaction of GAPR-1 with lipid bilayers is regulated by alternative homodimerization. Biochim Biophys Acta 1818: 2175–2183. |
| [18] | Olrichs NK, Mahalka AK, Kaloyanova D, et al. (2014) Golgi-Associated plant Pathogenesis Related protein 1 (GAPR-1) forms amyloid-like fibrils by interaction with acidic phospholipids and inhibits Abeta aggregation. Amyloid 21: 88–96. |
| [19] | Yoshiike Y, Minai R, Matsuo Y, et al. (2008) Amyloid oligomer conformation in a group of natively folded proteins. PLoS One 3: e3235. |
| [20] | Schwarzman AL, Gregori L, Vitek MP, et al. (1994) Transthyretin sequesters amyloid beta protein and prevents amyloid formation. Proc Natl Acad Sci USA 91: 8368–8372. |
| [21] | Chen J, Yagi H, Sormanni P, et al. (2012) Fibrillogenic propensity of the GroEL apical domain: a Janus-faced minichaperone. FEBS Lett 586: 1120–1127. |
| [22] | Rekas A, Ahn KJ, Kim J, et al. (2012) The chaperone activity of alpha-synuclein: Utilizing deletion mutants to map its interaction with target proteins. Proteins 80: 1316–1325. |
| [23] | Ofengeim D, Ito Y, Najafov A, et al. (2015) Activation of necroptosis in multiple sclerosis. Cell Rep 10: 1836–1849. |
| [24] | Zhang L, Qu S, Liang A, et al. (2015) Gene expression microarray analysis of the sciatic nerve of mice with diabetic neuropathy. Int J Mol Med 35: 333–339. |
| [25] | Karlsson O, Berg AL, Hanrieder J, et al. (2015) Intracellular fibril formation, calcification, and enrichment of chaperones, cytoskeletal, and intermediate filament proteins in the adult hippocampus CA1 following neonatal exposure to the nonprotein amino acid BMAA. Arch Toxicol 89: 423–436. |
| [26] | Nishi H, Hashimoto K, Madej T, et al. (2013) Evolutionary, physicochemical, and functional mechanisms of protein homooligomerization. Prog Mol Biol Transl Sci 117: 3–24. |
| [27] | Marianayagam NJ, Sunde M, Matthews JM (2004) The power of two: protein dimerization in biology. Trends Biochem Sci 29: 618–625. |
| [28] | Ali MH, Imperiali B (2005) Protein oligomerization: how and why. Bioorg Med Chem 13: 5013–5020. |
| [29] | Wang X, Sharp JS, Handel TM, et al. (2013) Chemokine oligomerization in cell signaling and migration. Prog Mol Biol Transl Sci 117: 531–578. |
| [30] | Zimmerberg J, Kozlov MM (2006) How proteins produce cellular membrane curvature. Nat Rev Mol Cell Biol 7: 9–19. |
| [31] | Park PS, Filipek S, Wells JW, et al. (2004) Oligomerization of G protein-coupled receptors: past, present, and future. Biochemistry 43: 15643–15656. |
| [32] | Adi-Harel S, Erlich S, Schmukler E, et al. (2010) Beclin 1 self-association is independent of autophagy induction by amino acid deprivation and rapamycin treatment. J Cell Biochem 110: 1262–1271. |
| [33] | Ku B, Woo JS, Liang C, et al. (2008) An insight into the mechanistic role of Beclin 1 and its inhibition by prosurvival Bcl-2 family proteins. Autophagy 4: 519–520. |
| [34] | Chiti F, Dobson CM (2006) Protein misfolding, functional amyloid, and human disease. Annu Rev Biochem 75: 333–366. |
| [35] | Dobson CM (2003) Protein folding and misfolding. Nature 426: 884–890. |
| [36] | Iannuzzi C, Irace G, Sirangelo I (2015) The effect of glycosaminoglycans (GAGs) on amyloid aggregation and toxicity. Molecules 20: 2510–2528. |
| [37] | Oskarsson ME, Singh K, Wang J, et al. (2015) Heparan Sulfate Proteoglycans Are Important for Islet Amyloid Formation and Islet Amyloid Polypeptide-induced Apoptosis. J Biol Chem 290: 15121–15132. |
| [38] | Kinnunen PK (2009) Amyloid Formation on Lipid Membrane Surfaces. The Open Biology Journal 2: 163–175. |
| [39] | Burke KA, Yates EA, Legleiter J (2013) Biophysical insights into how surfaces, including lipid membranes, modulate protein aggregation related to neurodegeneration. Front Neurol 4: 17. |
| [40] | Sunde M, Serpell LC, Bartlam M, et al. (1997) Common core structure of amyloid fibrils by synchrotron X-ray diffraction. J Mol Biol 273: 729–739. |
| [41] | Glabe CG (2008) Structural classification of toxic amyloid oligomers. J Biol Chem 283: 29639–29643. |
| [42] | Kayed R, Lasagna-Reeves CA (2013) Molecular mechanisms of amyloid oligomers toxicity. J Alzheimers Dis 33 Suppl 1: S67–78. |
| [43] | Breydo L, Uversky VN (2015) Structural, morphological, and functional diversity of amyloid oligomers. FEBS Lett 589: 2640–2648. |
| [44] | Haass C, Selkoe DJ (2007) Soluble protein oligomers in neurodegeneration: lessons from the Alzheimer's amyloid beta-peptide. Nat Rev Mol Cell Biol 8: 101–112. |
| [45] | Kagan BL, Jang H, Capone R, et al. (2012) Antimicrobial properties of amyloid peptides. Mol Pharm 9: 708–717. |
| [46] | Greenwald J, Riek R (2012) On the possible amyloid origin of protein folds. J Mol Biol 421: 417–426. |
| [47] | Gsponer J, Babu MM (2012) Cellular strategies for regulating functional and nonfunctional protein aggregation. Cell Rep 2: 1425–1437. |
| [48] | Fowler DM, Koulov AV, Balch WE, et al. (2007) Functional amyloid--from bacteria to humans. Trends Biochem Sci 32: 217–224. |
| [49] | Maji SK, Perrin MH, Sawaya MR, et al. (2009) Functional amyloids as natural storage of peptide hormones in pituitary secretory granules. Science 325: 328–332. |
| [50] | Puzzo D, Arancio O (2013) Amyloid-beta peptide: Dr. Jekyll or Mr. Hyde? J Alzheimers Dis 33 Suppl 1: S111–20. |
| [51] | Hervas R, Li L, Majumdar A, et al. (2016) Molecular Basis of Orb2 Amyloidogenesis and Blockade of Memory Consolidation. PLoS Biol 14: e1002361. |
| [52] | Jacob RS, George E, Singh PK, et al. (2016) Cell Adhesion on Amyloid Fibrils Lacking Integrin Recognition Motif. J Biol Chem . |
| [53] | Halfmann R, Jarosz DF, Jones SK, et al. (2012) Prions are a common mechanism for phenotypic inheritance in wild yeasts. Nature 482: 363–368. |
| [54] | Li J, McQuade T, Siemer AB, et al. (2012) The RIP1/RIP3 necrosome forms a functional amyloid signaling complex required for programmed necrosis. Cell 150: 339–350. |
| [55] | Parry TL, Melehani JH, Ranek MJ, et al. (2015) Functional Amyloid Signaling via the Inflammasome, Necrosome, and Signalosome: New Therapeutic Targets in Heart Failure. Front Cardiovasc Med 2: 25. |
| [56] | Rapsinski GJ, Wynosky-Dolfi MA, Oppong GO, et al. (2015) Toll-like receptor 2 and NLRP3 cooperate to recognize a functional bacterial amyloid, curli. Infect Immun 83: 693–701. |
| [57] | Heneka MT, Kummer MP, Latz E (2014) Innate immune activation in neurodegenerative disease. Nat Rev Immunol 14: 463–477. |
| [58] | Maldera JA, Vasen G, Ernesto JI, et al. (2011) Evidence for the involvement of zinc in the association of CRISP1 with rat sperm during epididymal maturation. Biol Reprod 85: 503–510. |
| [59] | Lu S, Faris JD, Sherwood R, et al. (2014) A dimeric PR-1-type pathogenesis-related protein interacts with ToxA and potentially mediates ToxA-induced necrosis in sensitive wheat. Mol Plant Pathol 15: 650–663. |
| [60] | Borloo J, Geldhof P, Peelaers I, et al. (2013) Structure of Ostertagia ostertagi ASP-1: insights into disulfide-mediated cyclization and dimerization. Acta Crystallogr D Biol Crystallogr 69: 493–503. |
| [61] | Prados-Rosales RC, Roldan-Rodriguez R, Serena C, et al. (2012) A PR-1-like protein of Fusarium oxysporum functions in virulence on mammalian hosts. J Biol Chem 287: 21970–21979. |
| [62] | Sugiyama H, Burnett L, Xiang X, et al. (2009) Purification and multimer formation of allurin, a sperm chemoattractant from Xenopus laevis egg jelly. Mol Reprod Dev 76: 527–536. |
| [63] | Asojo OA, Goud G, Dhar K, et al. (2005) X-ray structure of Na-ASP-2, a pathogenesis-related-1 protein from the nematode parasite, Necator americanus, and a vaccine antigen for human hookworm infection. J Mol Biol 346: 801–814. |
| [64] | Ma D, Francischetti IM, Ribeiro JM, et al. (2015) The structure of hookworm platelet inhibitor (HPI), a CAP superfamily member from Ancylostoma caninum. Acta Crystallogr F Struct Biol Commun 71: 643–649. |
| [65] | Tsolis AC, Papandreou NC, Iconomidou VA, et al. (2013) A consensus method for the prediction of 'aggregation-prone' peptides in globular proteins. PLoS One 8: e54175. |
| [66] | Egge N, Muthusubramanian A, Cornwall GA (2015) Amyloid properties of the mouse egg zona pellucida. PLoS One 10: e0129907. |
| [67] | Guyonnet B, Egge N, Cornwall GA (2014) Functional amyloids in the mouse sperm acrosome. Mol Cell Biol 34: 2624–2634. |
| [68] | Swanson WJ, Aagaard JE, Vacquier VD, et al. (2011) The molecular basis of sex: linking yeast to human. Mol Biol Evol 28: 1963–1966. |
| [69] | Garcia MC, Lee JT, Ramsook CB, et al. (2011) A role for amyloid in cell aggregation and biofilm formation. PLoS One 6: e17632. |
| [70] | Da Ros VG, Munoz MW, Battistone MA, et al. (2015) From the epididymis to the egg: participation of CRISP proteins in mammalian fertilization. Asian J Androl 17: 711–715. |
| [71] | Caballero J, Frenette G, D'Amours O, et al. (2012) Bovine sperm raft membrane associated Glioma Pathogenesis-Related 1-like protein 1 (GliPr1L1) is modified during the epididymal transit and is potentially involved in sperm binding to the zona pellucida. J Cell Physiol 227: 3876–3886. |
| [72] | Busso D, Goldweic NM, Hayashi M, et al. (2007) Evidence for the involvement of testicular protein CRISP2 in mouse sperm-egg fusion. Biol Reprod 76: 701–708. |
| [73] | Doty A, Buhi WC, Benson S, et al. (2011) Equine CRISP3 modulates interaction between spermatozoa and polymorphonuclear neutrophils. Biol Reprod 85: 157–164. |
| [74] | Ernesto JI, Weigel Munoz M, Battistone MA, et al. (2015) CRISP1 as a novel CatSper regulator that modulates sperm motility and orientation during fertilization. J Cell Biol 210: 1213–1224. |
| [75] | Burnett LA, Xiang X, Bieber AL, et al. (2008) Crisp proteins and sperm chemotaxis: discovery in amphibians and explorations in mammals. Int J Dev Biol 52: 489–501. |
| [76] | Burnett LA, Anderson DM, Rawls A, et al. (2011) Mouse sperm exhibit chemotaxis to allurin, a truncated member of the cysteine-rich secretory protein family. Dev Biol 360: 318–328. |
| [77] | Usmani SM, Zirafi O, Muller JA, et al. (2014) Direct visualization of HIV-enhancing endogenous amyloid fibrils in human semen. Nat Commun 5: 3508. |
| [78] | Castellano LM, Shorter J (2012) The Surprising Role of Amyloid Fibrils in HIV Infection. Biology (Basel) 1: 58–80. |
| [79] | Doncel GF (2006) Exploiting common targets in human fertilization and HIV infection: development of novel contraceptive microbicides. Hum Reprod Update 12: 103–117. |
| [80] | Gebbink MF, Claessen D, Bouma B, et al. (2005) Amyloids--a functional coat for microorganisms. Nat Rev Microbiol 3: 333–341. |
| [81] | Garcia-Sherman MC, Lundberg T, Sobonya RE, et al. (2015) A unique biofilm in human deep mycoses: fungal amyloid is bound by host serum amyloid P component. NPJ Biofilms Microbiomes 1: 15009. |
| [82] | Syed AK, Boles BR (2014) Fold modulating function: bacterial toxins to functional amyloids. Front Microbiol 5: 401. |
| [83] | Pisa D, Alonso R, Rabano A, et al. (2015) Different Brain Regions are Infected with Fungi in Alzheimer's Disease. Sci Rep 5: 15015. |
| [84] | Oh J, Kim JG, Jeon E, et al. (2007) Amyloidogenesis of type III-dependent harpins from plant pathogenic bacteria. J Biol Chem 282: 13601–13609. |
| [85] | van Loon LC, Rep M, Pieterse CM (2006) Significance of inducible defense-related proteins in infected plants. Annu Rev Phytopathol 44: 135–162. |
| [86] | Van Loon LC, Van Strien EA (1999) The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol Mol Plant Pathol 55: 85–97. |
| [87] | Niderman T, Genetet I, Bruyere T, et al. (1995) Pathogenesis-related PR-1 proteins are antifungal. Isolation and characterization of three 14-kilodalton proteins of tomato and of a basic PR-1 of tobacco with inhibitory activity against Phytophthora infestans. Plant Physiol 108: 17–27. |
| [88] | Rauscher M, Adam AL, Wirtz S, et al. (1999) PR-1 protein inhibits the differentiation of rust infection hyphae in leaves of acquired resistant broad bean. Plant J 19: 625–633. |
| [89] | Benhamou N (1995) Immunocytochemistry of plant defense mechanisms induced upon microbial attack. Microsc Res Tech 31: 63–78. |
| [90] | Lee YK, Hippe-Sanwald S, Lee SC, et al. (2000) In situ localization of PR-1 mRNA and PR-1 protein in compatible and incompatible interactions of pepper stems with Phytophthora capsici. Protoplasma 211: 64–75. |
| [91] | Santén K, Marttila S, Liljeroth E, et al. (2005) Immunocytochemical localization of the pathogenesis-related PR-1 protein in barley leaves after infection by Bipolaris sorokiniana. Physiol Mol Plant Pathol 66: 45–54. |
| [92] | Teixeira PJ, Costa GG, Fiorin GL, et al. (2013) Novel receptor-like kinases in cacao contain PR-1 extracellular domains. Mol Plant Pathol 14: 602–609. |
| [93] | Jiang G, Hunter T (1999) Receptor signaling: when dimerization is not enough. Curr Biol 9: R568–71. |
| [94] | Chen YL, Lee CY, Cheng KT, et al. (2014) Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato. Plant Cell 26: 4135–4148. |
| [95] | Chien PS, Nam HG, Chen YR (2015) A salt-regulated peptide derived from the CAP superfamily protein negatively regulates salt-stress tolerance in Arabidopsis. J Exp Bot 66: 5301–5313. |
| [96] | Buch F, Pauchet Y, Rott M, et al. (2014) Characterization and heterologous expression of a PR-1 protein from traps of the carnivorous plant Nepenthes mirabilis. Phytochemistry 100: 43–50. |
| [97] | Osman A, Wang CK, Winter A, et al. (2012) Hookworm SCP/TAPS protein structure--A key to understanding host-parasite interactions and developing new interventions. Biotechnol Adv 30: 652–657. |
| [98] | Moyle M, Foster DL, McGrath DE, et al. (1994) A hookworm glycoprotein that inhibits neutrophil function is a ligand of the integrin CD11b/CD18. J Biol Chem 269: 10008–10015. |
| [99] | Asojo OA (2011) Structure of a two-CAP-domain protein from the human hookworm parasite Necator americanus. Acta Crystallogr D Biol Crystallogr 67: 455–462. |
| [100] | Del Valle A, Jones BF, Harrison LM, et al. (2003) Isolation and molecular cloning of a secreted hookworm platelet inhibitor from adult Ancylostoma caninum. Mol Biochem Parasitol 129: 167–177. |
| [101] | Horn M, Bertling A, Brodde MF, et al. (2012) Human neutrophil alpha-defensins induce formation of fibrinogen and thrombospondin-1 amyloid-like structures and activate platelets via glycoprotein IIb/IIIa. J Thromb Haemost 10: 647–661. |
| [102] | Canobbio I, Abubaker AA, Visconte C, et al. (2015) Role of amyloid peptides in vascular dysfunction and platelet dysregulation in Alzheimer's disease. Front Cell Neurosci 9: 65. |
| [103] | Wang YL, Kuo JH, Lee SC, et al. (2010) Cobra CRISP functions as an inflammatory modulator via a novel Zn2+- and heparan sulfate-dependent transcriptional regulation of endothelial cell adhesion molecules. J Biol Chem 285: 37872–37883. |
| [104] | Alexandrescu AT (2005) Amyloid accomplices and enforcers. Protein Sci 14: 1–12. |
| [105] | Hughes AJ, Hussain R, Cosentino C, et al. (2012) A zinc complex of heparan sulfate destabilises lysozyme and alters its conformation. Biochem Biophys Res Commun 425: 794–799. |
| [106] | Berthelot K, Lecomte S, Coulary-Salin B, et al. (2016) Hevea brasiliensis prohevein possesses a conserved C-terminal domain with amyloid-like properties in vitro. Biochim Biophys Acta 1864: 388–399. |
| [107] | Lay FT, Mills GD, Poon IK, et al. (2012) Dimerization of plant defensin NaD1 enhances its antifungal activity. J Biol Chem 287: 19961–19972. |
| [108] | Poon IK, Baxter AA, Lay FT, et al. (2014) Phosphoinositide-mediated oligomerization of a defensin induces cell lysis. Elife 3: e01808. |
| [109] | Garvey M, Meehan S, Gras SL, et al. (2013) A radish seed antifungal peptide with a high amyloid fibril-forming propensity. Biochim Biophys Acta 1834: 1615–1623. |
| [110] | Marangon M, Van Sluyter SC, Waters EJ, et al. (2014) Structure of haze forming proteins in white wines: Vitis vinifera thaumatin-like proteins. PLoS One 9: e113757. |
| [111] | Yu XM, Griffith M (1999) Antifreeze proteins in winter rye leaves form oligomeric complexes. Plant Physiol 119: 1361–1370. |
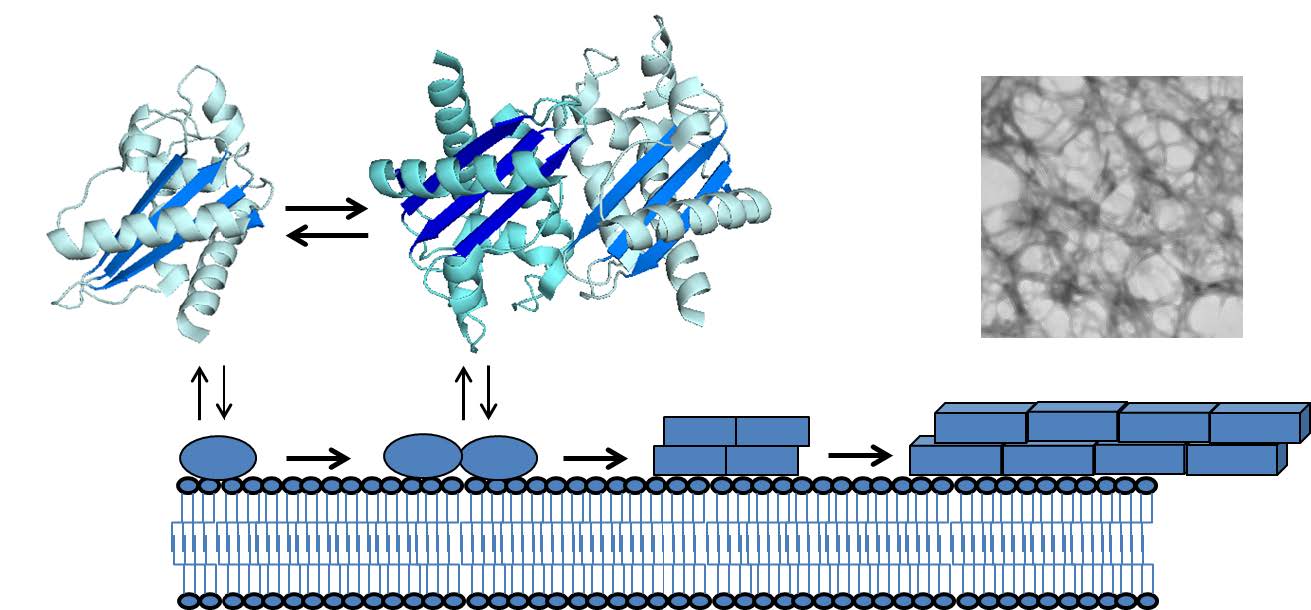

Figures(2)

Nick K. Olrichs, J. Bernd Helms. Novel insights into the function of the conserved domain of the CAP superfamily of proteins[J]. AIMS Biophysics, 2016, 3(2): 232-246. doi: 10.3934/biophy.2016.2.232







 DownLoad:
DownLoad: