To investigate the properties of a large-scale brain network, it is a common practice to reduce the dimension of resting state functional magnetic resonance imaging (rs-fMRI) data to tens to hundreds of nodes. This study presents an analytic streamline that incorporates modular analysis and similarity measurements (MOSI) to fulfill functional parcellation (FP) of the cortex. MOSI is carried out by iteratively dividing a module into sub-modules (via the Louvain community detection method) and unifying similar neighboring sub-modules into a new module (adjacent sub-modules with a similarity index <0.05) until the brain modular structures of successive runs become constant. By adjusting the gamma value, a parameter in the Louvain algorithm, MOSI may segment the cortex with different resolutions. rs-fMRI scans of 33 healthy subjects were selected from the dataset of the Rockland sample. MOSI was applied to the rs-fMRI data after standardized pre-processing steps. The results indicate that the parcellated modules by MOSI are more homogeneous in content. After reducing the grouped voxels to representative neural nodes, the network structures were explored. The resultant network components were comparable with previous reports. The validity of MOSI in achieving data reduction has been confirmed. MOSI may provide a novel starting point for further investigation of the network properties of rs-fMRI data. Potential applications of MOSI are discussed.
Citation: Tien-Wen Lee, Gerald Tramontano. Automatic parcellation of resting-state cortical dynamics by iterative community detection and similarity measurements[J]. AIMS Neuroscience, 2021, 8(4): 526-542. doi: 10.3934/Neuroscience.2021028
To investigate the properties of a large-scale brain network, it is a common practice to reduce the dimension of resting state functional magnetic resonance imaging (rs-fMRI) data to tens to hundreds of nodes. This study presents an analytic streamline that incorporates modular analysis and similarity measurements (MOSI) to fulfill functional parcellation (FP) of the cortex. MOSI is carried out by iteratively dividing a module into sub-modules (via the Louvain community detection method) and unifying similar neighboring sub-modules into a new module (adjacent sub-modules with a similarity index <0.05) until the brain modular structures of successive runs become constant. By adjusting the gamma value, a parameter in the Louvain algorithm, MOSI may segment the cortex with different resolutions. rs-fMRI scans of 33 healthy subjects were selected from the dataset of the Rockland sample. MOSI was applied to the rs-fMRI data after standardized pre-processing steps. The results indicate that the parcellated modules by MOSI are more homogeneous in content. After reducing the grouped voxels to representative neural nodes, the network structures were explored. The resultant network components were comparable with previous reports. The validity of MOSI in achieving data reduction has been confirmed. MOSI may provide a novel starting point for further investigation of the network properties of rs-fMRI data. Potential applications of MOSI are discussed.

| [1] |
Lee TW, Xue SW (2017) Linking graph features of anatomical architecture to regional brain activity: A multi-modal mri study. Neurosci. Lett 651: 123-127. doi: 10.1016/j.neulet.2017.05.005

|
| [2] |
Worsley KJ, Taylor JE, Tomaiuolo F, et al. (2004) Unified univariate and multivariate random field theory. Neuroimage 23: S189-S195. doi: 10.1016/j.neuroimage.2004.07.026
|
| [3] |
Cox RW (1996) Afni: Software for analysis and visualization of functional magnetic resonance neuroimages. Comput Biomed Res 29: 162-173. doi: 10.1006/cbmr.1996.0014
|
| [4] |
Xue SW, Li D, Weng XC, et al. (2014) Different neural manifestations of two slow frequency bands in resting functional magnetic resonance imaging: A systemic survey at regional, interregional, and network levels. Brain Connect 4: 242-255. doi: 10.1089/brain.2013.0182
|
| [5] |
Lee TW, Northoff G, Wu YT (2014) Resting network is composed of more than one neural pattern: An fmri study. Neuroscience 274: 198-208. doi: 10.1016/j.neuroscience.2014.05.035
|
| [6] |
Lee TW, Xue SW (2017) Examination of the validity of the atlas-informed approach to functional parcellation: A resting functional mri study. Neuroreport 28: 649-653. doi: 10.1097/WNR.0000000000000808
|
| [7] |
Raichle ME, Snyder AZ (2007) A default mode of brain function: A brief history of an evolving idea. Neuroimage 37: 1083-1090. doi: 10.1016/j.neuroimage.2007.02.041
|
| [8] |
Greicius MD, Supekar K, Menon V, et al. (2009) Resting-state functional connectivity reflects structural connectivity in the default mode network. Cereb Cortex 19: 72-78. doi: 10.1093/cercor/bhn059
|
| [9] |
Calhoun VD, Adali T (2012) Multisubject independent component analysis of fmri: A decade of intrinsic networks, default mode, and neurodiagnostic discovery. IEEE Rev Biomed Eng 5: 60-73. doi: 10.1109/RBME.2012.2211076
|
| [10] |
Amunts K, Schleicher A, Burgel U, et al. (1999) Broca's region revisited: Cytoarchitecture and intersubject variability. J Comp Neurol 412: 319-341. doi: 10.1002/(SICI)1096-9861(19990920)412:2<319::AID-CNE10>3.0.CO;2-7
|
| [11] |
Wig GS, Schlaggar BL, Petersen SE (2011) Concepts and principles in the analysis of brain networks. Ann N Y Acad Sci 1224: 126-146. doi: 10.1111/j.1749-6632.2010.05947.x
|
| [12] |
Smith SM, Miller KL, Salimi-Khorshidi G, et al. (2011) Network modelling methods for fmri. Neuroimage 54: 875-891. doi: 10.1016/j.neuroimage.2010.08.063
|
| [13] |
Stanley ML, Moussa MN, Paolini BM, Lyday, et al. (2013) Defining nodes in complex brain networks. Front Comput Neurosci 7: 169. doi: 10.3389/fncom.2013.00169
|
| [14] |
Hilgetag CC, O'Neill MA, Young MP (2000) Hierarchical organization of macaque and cat cortical sensory systems explored with a novel network processor. Philos Trans R Soc Lond B Biol Sci 355: 71-89. doi: 10.1098/rstb.2000.0550
|
| [15] |
Felleman DJ, Van Essen DC (1991) Distributed hierarchical processing in the primate cerebral cortex. Cereb Cortex 1: 1-47. doi: 10.1093/cercor/1.1.1
|
| [16] |
Van Essen DC, Anderson CH, Felleman DJ (1992) Information processing in the primate visual system: An integrated systems perspective. Science 255: 419-423. doi: 10.1126/science.1734518
|
| [17] |
Lee TW, Xue SW (2018) Does emotion regulation engage the same neural circuit as working memory? A meta-analytical comparison between cognitive reappraisal of negative emotion and 2-back working memory task. PLoS One 13: e0203753. doi: 10.1371/journal.pone.0203753
|
| [18] |
Lee TW, Dolan RJ, Critchley HD (2008) Controlling emotional expression: Behavioral and neural correlates of nonimitative emotional responses. Cereb Cortex 18: 104-113. doi: 10.1093/cercor/bhm035
|
| [19] |
Lee TW, Josephs O, Dolan RJ, et al. (2006) Imitating expressions: Emotion-specific neural substrates in facial mimicry. Soc Cogn Affect Neurosci 1: 122-135. doi: 10.1093/scan/nsl012
|
| [20] |
Liakakis G, Nickel J, Seitz RJ (2011) Diversity of the inferior frontal gyrus--a meta-analysis of neuroimaging studies. Behav Brain Res 225: 341-347. doi: 10.1016/j.bbr.2011.06.022
|
| [21] |
Zhong YM, Rockland KS (2003) Inferior parietal lobule projections to anterior inferotemporal cortex (area te) in macaque monkey. Cereb Cortex 13: 527-540. doi: 10.1093/cercor/13.5.527
|
| [22] |
Hutchinson JB, Uncapher MR, Wagner AD (2009) Posterior parietal cortex and episodic retrieval: Convergent and divergent effects of attention and memory. Learn Mem 16: 343-356. doi: 10.1101/lm.919109
|
| [23] |
Schultz H, Sommer T, Peters J (2012) Direct evidence for domain-sensitive functional subregions in human entorhinal cortex. J Neurosci 32: 4716-4723. doi: 10.1523/JNEUROSCI.5126-11.2012
|
| [24] |
Winston JS, O'Doherty J, Dolan RJ (2003) Common and distinct neural responses during direct and incidental processing of multiple facial emotions. Neuroimage 20: 84-97. doi: 10.1016/S1053-8119(03)00303-3
|
| [25] |
Grill-Spector K, Malach R (2004) The human visual cortex. Annu Rev Neurosci 27: 649-677. doi: 10.1146/annurev.neuro.27.070203.144220
|
| [26] |
Connolly AC, Guntupalli JS, Gors J, et al. (2012) The representation of biological classes in the human brain. J Neurosci 32: 2608-2618. doi: 10.1523/JNEUROSCI.5547-11.2012
|
| [27] |
Astafiev SV, Stanley CM, Shulman GL, et al. (2001) Extrastriate body area in human occipital cortex responds to the performance of motor actions. Nat Neurosci 7: 542-548. doi: 10.1038/nn1241
|
| [28] |
Yeo BT, Krienen FM, Sepulcre J, et al. (2011) The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J Neurophysiol 106: 1125-1165. doi: 10.1152/jn.00338.2011
|
| [29] |
Hilgetag CC, Burns GA, O'Neill MA, et al. (2000) Anatomical connectivity defines the organization of clusters of cortical areas in the macaque monkey and the cat. Philos Trans R Soc Lond B Biol Sci 355: 91-110. doi: 10.1098/rstb.2000.0551
|
| [30] |
Moussa MN, Steen MR, Laurienti PJ, et al. (2012) Consistency of network modules in resting-state fmri connectome data. PLoS One 7: e44428. doi: 10.1371/journal.pone.0044428
|
| [31] |
Cohen AL, Fair DA, Dosenbach NU, et al. (2008) Defining functional areas in individual human brains using resting functional connectivity mri. Neuroimage 41: 45-57. doi: 10.1016/j.neuroimage.2008.01.066
|
| [32] |
Shen X, Papademetris X, Constable RT (2010) Graph-theory based parcellation of functional subunits in the brain from resting-state fmri data. Neuroimage 50: 1027-1035. doi: 10.1016/j.neuroimage.2009.12.119
|
| [33] |
Ryali S, Chen T, Supekar K, et al. (2013) A parcellation scheme based on von mises-fisher distributions and markov random fields for segmenting brain regions using resting-state fmri. Neuroimage 65: 83-96. doi: 10.1016/j.neuroimage.2012.09.067
|
| [34] |
Wig GS, Laumann TO, Cohen AL, et al. (2014) Parcellating an individual subject's cortical and subcortical brain structures using snowball sampling of resting-state correlations. Cereb Cortex 24: 2036-2054. doi: 10.1093/cercor/bht056
|
| [35] |
Wang D, Buckner RL, Fox MD, et al. (2015) Parcellating cortical functional networks in individuals. Nat Neurosci 18: 1853-1860. doi: 10.1038/nn.4164
|
| [36] |
Blondel VD, Guillaume JL, Lambiotte R, et al. (2008) Fast unfolding of communities in large networks. J Stat Mech Theory Exp 2008: P10008. doi: 10.1088/1742-5468/2008/10/P10008
|
| [37] |
Rubinov M, Sporns O (2011) Weight-conserving characterization of complex functional brain networks. Neuroimage 56: 2068-2079. doi: 10.1016/j.neuroimage.2011.03.069
|
| [38] |
Leicht EA, Newman ME (2008) Community structure in directed networks. Phys Rev Lett 100: 118703. doi: 10.1103/PhysRevLett.100.118703
|
| [39] |
Lee TW, Xue SW (2018) Functional connectivity maps based on hippocampal and thalamic dynamics may account for the default-mode network. Eur J Neurosci 47: 388-398. doi: 10.1111/ejn.13828
|
| [40] |
Dale AM, Fischl B, Sereno MI (1999) Cortical surface-based analysis. I. Segmentation and surface reconstruction. Neuroimage 9: 179-194. doi: 10.1006/nimg.1998.0395
|
| [41] |
Fischl B, Sereno MI, Dale AM (1999) Cortical surface-based analysis. Ii: Inflation, flattening, and a surface-based coordinate system. Neuroimage 9: 195-207. doi: 10.1006/nimg.1998.0396
|
| [42] |
Jo HJ, Saad ZS, Simmons WK, et al. (2010) Mapping sources of correlation in resting state fmri, with artifact detection and removal. Neuroimage 52: 571-582. doi: 10.1016/j.neuroimage.2010.04.246
|
| [43] |
Mucha PJ, Richardson T, Macon K, et al. (2010) Community structure in time-dependent, multiscale, and multiplex networks. Science 328: 876-878. doi: 10.1126/science.1184819
|
| [44] |
Reichardt J, Bornholdt S (2006) Statistical mechanics of community detection. Phys Rev E Stat Nonlin Soft Matter Phys 74: 016110. doi: 10.1103/PhysRevE.74.016110
|
| [45] |
Lee TW, Xue SW (2018) Revisiting the functional and structural connectivity of large-scale cortical networks. Brain Connect 8: 129-138. doi: 10.1089/brain.2017.0536
|
| [46] |
Samara Z, Evers EA, Goulas A, et al. (2017) Human orbital and anterior medial prefrontal cortex: Intrinsic connectivity parcellation and functional organization. Brain Struct Funct 222: 2941-2960. doi: 10.1007/s00429-017-1378-2
|
| [47] |
Goulas A, Uylings HB, Stiers P (2012) Unravelling the intrinsic functional organization of the human lateral frontal cortex: A parcellation scheme based on resting state fmri. J Neurosci 32: 10238-10252. doi: 10.1523/JNEUROSCI.5852-11.2012
|
| [48] |
Uddin LQ, Yeo BTT, Spreng RN (2019) Towards a universal taxonomy of macro-scale functional human brain networks. Brain Topogr 32: 926-942. doi: 10.1007/s10548-019-00744-6
|
| [49] | Golland P, Golland Y, Malach R (2007) Detection of spatial activation patterns as unsupervised segmentation of fmri data. Med Image Comput Comput Assist Interv 10: 110-118. |
| [50] | Tucholka A, Thirion B, Perrot M, et al. (2008) Probabilistic anatomo-functional parcellation of the cortex: How many regions? Med Image Comput Comput Assist Interv 11: 399-406. |
| [51] |
Kahnt T, Chang LJ, Park SQ, et al. (2012) Connectivity-based parcellation of the human orbitofrontal cortex. J Neurosci 32: 6240-6250. doi: 10.1523/JNEUROSCI.0257-12.2012
|
| [52] |
Craddock RC, James GA, Holtzheimer PE, et al. (2012) A whole brain fmri atlas generated via spatially constrained spectral clustering. Hum Brain Mapp 33: 1914-1928. doi: 10.1002/hbm.21333
|
| [53] | Liu J, Bai J, Peng M, et al. (2005) Analysis of functional mri data during continuous rest state sequently using time series and spatial clustering: A primary study. Conf Proc IEEE Eng Med Biol Soc 2005: 5291-5294. |
| [54] |
Kiviniemi V, Starck T, Remes J, et al. (2009) Functional segmentation of the brain cortex using high model order group pica. Hum Brain Mapp 30: 3865-3886. doi: 10.1002/hbm.20813
|
| [55] |
Varoquaux G, Sadaghiani S, Pinel P, et al. (2010) A group model for stable multi-subject ica on fmri datasets. Neuroimage 51: 288-299. doi: 10.1016/j.neuroimage.2010.02.010
|
| [56] |
Churchland PS, Sejnowski TJ (1988) Perspectives on cognitive neuroscience. Science 242: 741-745. doi: 10.1126/science.3055294
|
| [57] | Sejnowski TJ, Churchland PS (1989) Brain and cognition, foundations of cognitive science Cambridge, MA: MIT Press. |
| [58] |
Biswal B, Yetkin FZ, Haughton VM, et al. (1995) Functional connectivity in the motor cortex of resting human brain using echo-planar mri. Magn Reson Med 34: 537-541. doi: 10.1002/mrm.1910340409
|
| [59] |
Bassett DS, Wymbs NF, Porter MA, et al. (2011) Dynamic reconfiguration of human brain networks during learning. Proc Natl Acad Sci U S A 108: 7641-7646. doi: 10.1073/pnas.1018985108
|
| [60] |
Mega MS, Cummings JL, Salloway S, et al. (1997) The limbic system: An anatomic, phylogenetic, and clinical perspective. J Neuropsychiatry Clin Neurosci 9: 315-330. doi: 10.1176/jnp.9.3.315
|
| [61] |
Mazziotta J, Toga A, Evans A, et al. (2001) A probabilistic atlas and reference system for the human brain: International consortium for brain mapping (icbm). Philos Trans R Soc Lond B Biol Sci 356: 1293-1322. doi: 10.1098/rstb.2001.0915
|
| [62] |
Desikan RS, Segonne F, Fischl B, et al. (2006) An automated labeling system for subdividing the human cerebral cortex on mri scans into gyral based regions of interest. Neuroimage 31: 968-980. doi: 10.1016/j.neuroimage.2006.01.021
|
| [63] |
Aldecoa R, Marin I (2013) Surprise maximization reveals the community structure of complex networks. Sci Rep 3: 1060. doi: 10.1038/srep01060
|
| [64] |
Lancichinetti A, Radicchi F, Ramasco JJ, et al. (2011) Finding statistically significant communities in networks. PLoS One 6: e18961. doi: 10.1371/journal.pone.0018961
|
| [65] |
Good BH, de Montjoye YA, Clauset A (2010) Performance of modularity maximization in practical contexts. Phys Rev E 81: 046106. doi: 10.1103/PhysRevE.81.046106
|
| [66] |
Thirion B, Varoquaux G, Dohmatob E, et al. (2014) Which fmri clustering gives good brain parcellations? Front Neurosci 8: 167. doi: 10.3389/fnins.2014.00167
|
| [67] |
Duboc V, Dufourcq P, Blader P, et al. (2015) Asymmetry of the brain: Development and implications. Annu Rev Genet 49: 647-672. doi: 10.1146/annurev-genet-112414-055322
|
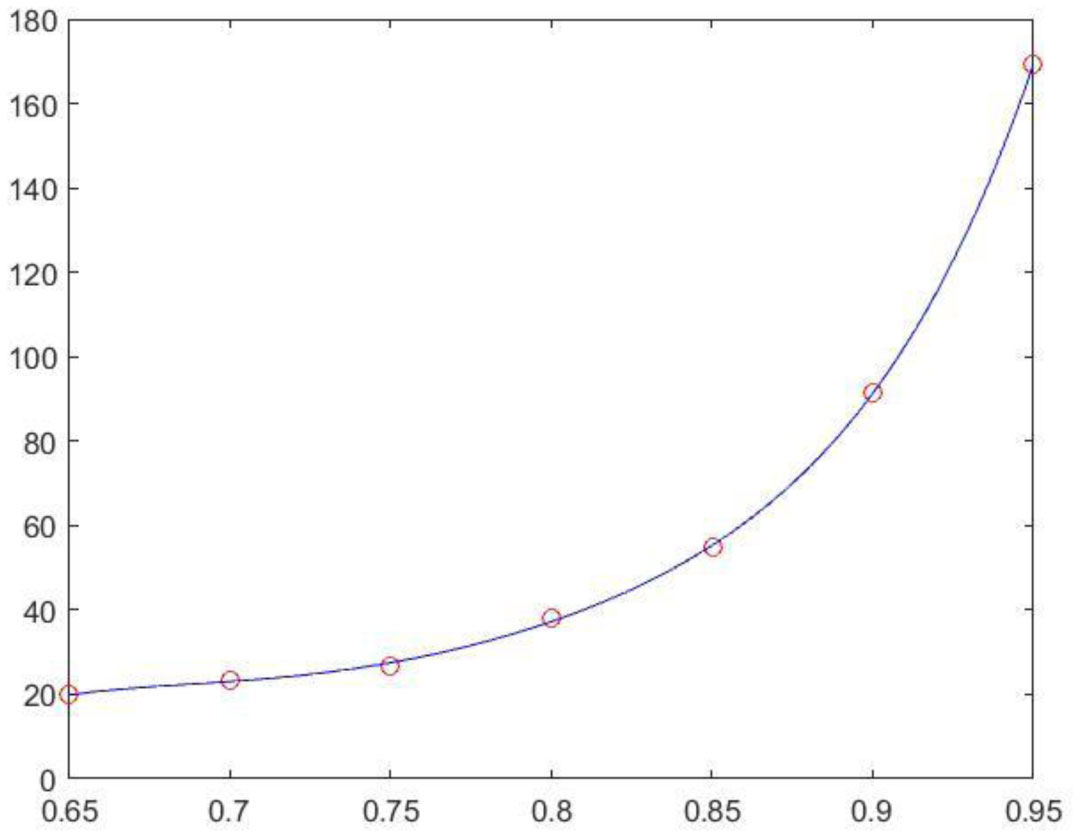
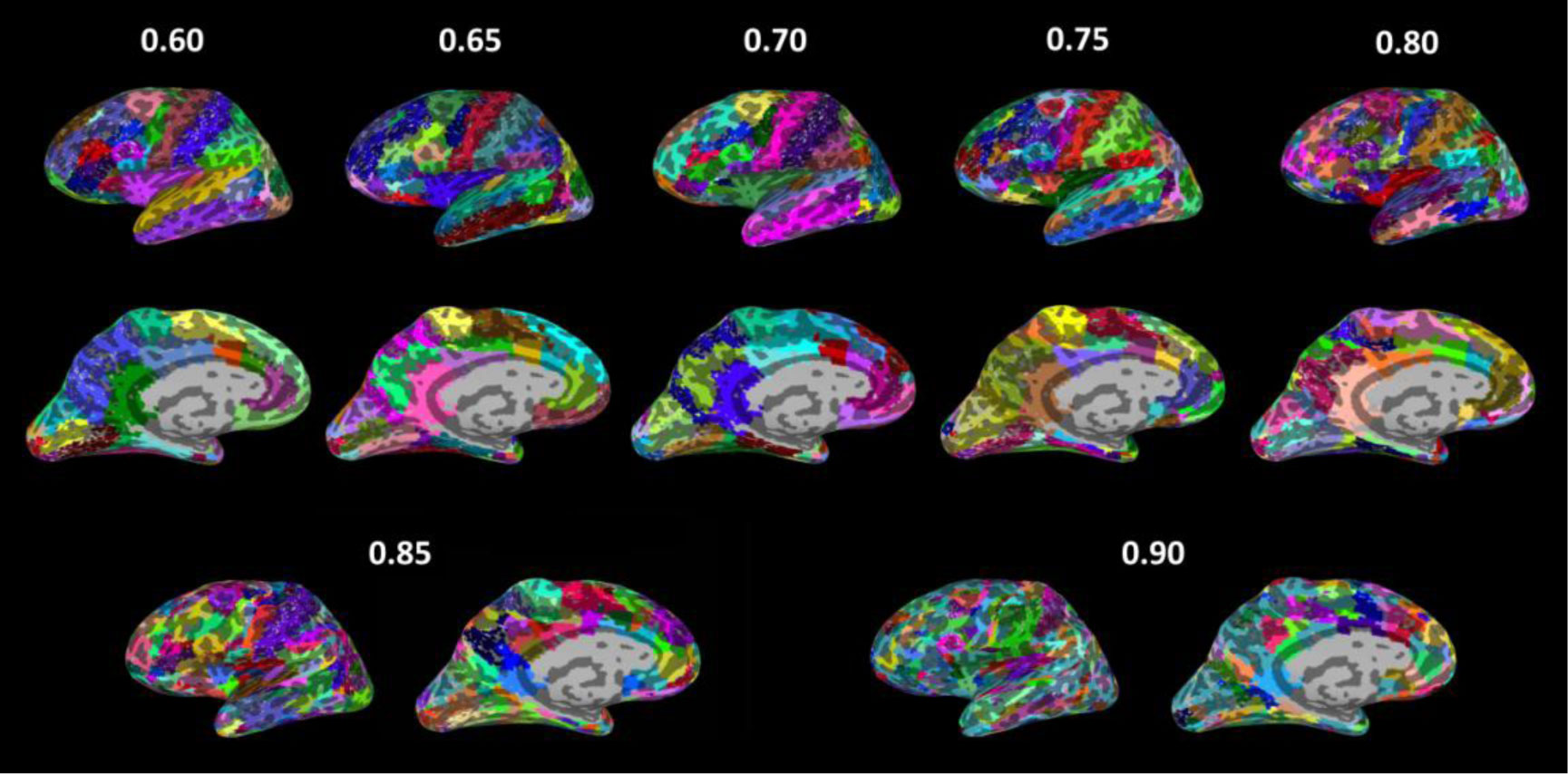
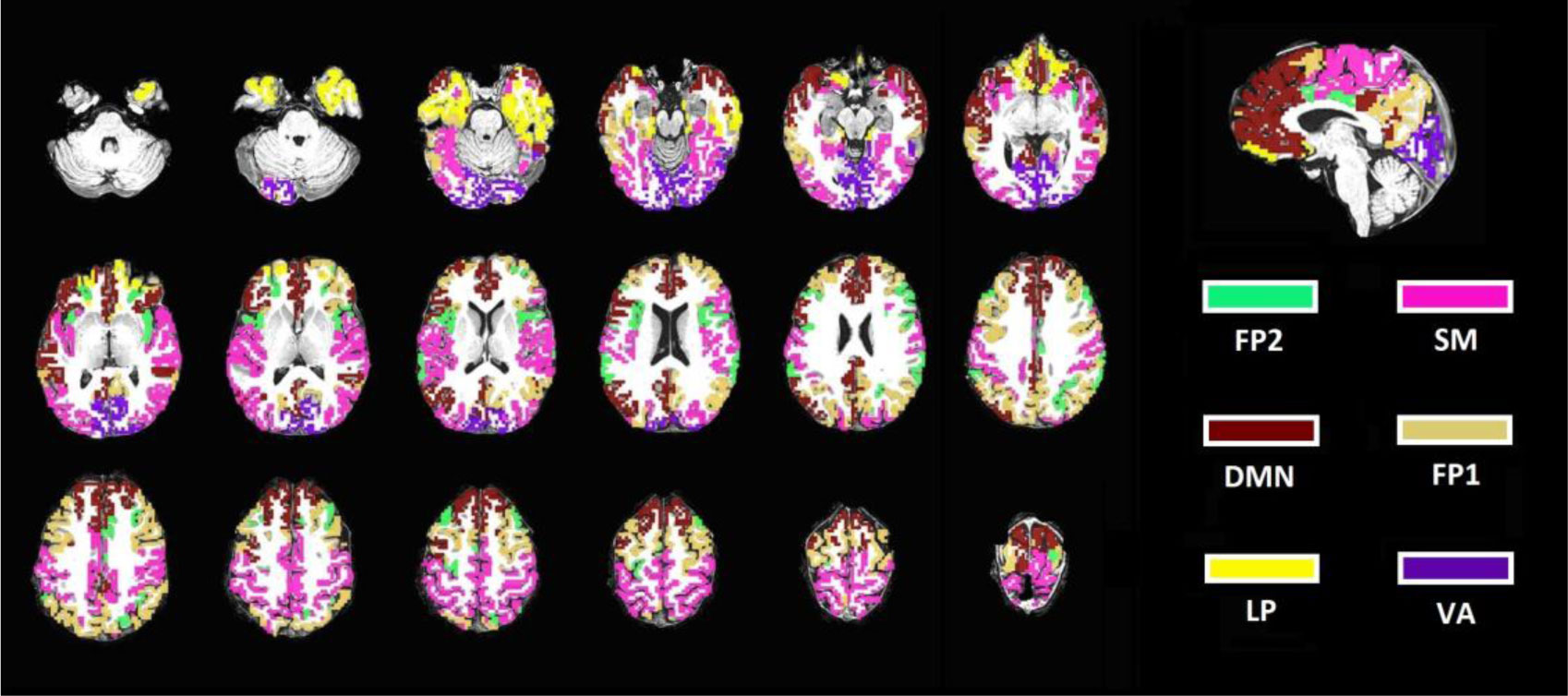
Figures(3) / Tables(3)

Tien-Wen Lee, Gerald Tramontano. Automatic parcellation of resting-state cortical dynamics by iterative community detection and similarity measurements[J]. AIMS Neuroscience, 2021, 8(4): 526-542. doi: 10.3934/Neuroscience.2021028







 DownLoad:
DownLoad: