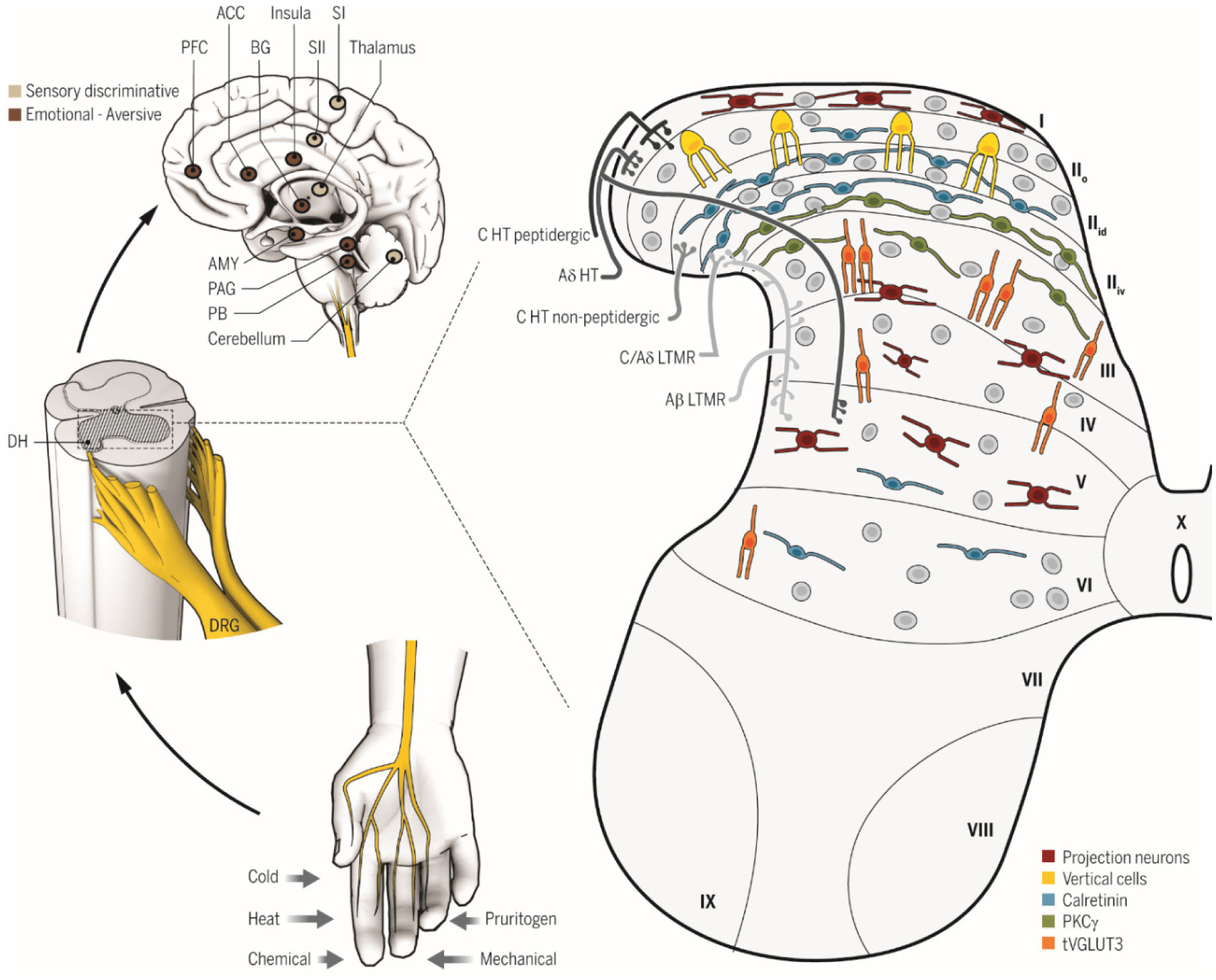
The current paper provides an explanation of neurophysiological pain processing based the Dimensional Systems Model (DSM), a theory of higher cortical functions in which the cortical column is considered the binary digit for all cortical functions. Within the discussion, novel views on the roles of the basal ganglia, cerebellum, and cingulate cortex are presented. Additionally, an applied Clinical Biopsychological Model (CBM) based on the DSM will be discussed as related to psychological treatment with chronic pain patients. Three specific areas that have not been adequately addressed in the psychological treatment of chronic pain patients will be discussed based on the CBM. The treatment approaches have been effectively used in a clinical setting. Conclusions focus on a call for researchers and clinicians to fully evaluate the value of both the DSM and CBM.
Citation: Robert A. Moss. Psychotherapy in pain management: New viewpoints and treatment targets based on a brain theory[J]. AIMS Neuroscience, 2020, 7(3): 194-207. doi: 10.3934/Neuroscience.2020013
The current paper provides an explanation of neurophysiological pain processing based the Dimensional Systems Model (DSM), a theory of higher cortical functions in which the cortical column is considered the binary digit for all cortical functions. Within the discussion, novel views on the roles of the basal ganglia, cerebellum, and cingulate cortex are presented. Additionally, an applied Clinical Biopsychological Model (CBM) based on the DSM will be discussed as related to psychological treatment with chronic pain patients. Three specific areas that have not been adequately addressed in the psychological treatment of chronic pain patients will be discussed based on the CBM. The treatment approaches have been effectively used in a clinical setting. Conclusions focus on a call for researchers and clinicians to fully evaluate the value of both the DSM and CBM.

| [1] |
Tseli E, Boersma K, Stålnacke BM, et al. (2019) Prognostic factors for physical functioning after multidisciplinary rehabilitation in patients with chronic musculoskeletal pain: a systematic review and meta-analysis. Clin J Pain 35: 148-173. doi: 10.1097/AJP.0000000000000669

|
| [2] |
Engel GL (1977) The need for a new medical model: a challenge for biomedicine. Science 196: 129-136. doi: 10.1126/science.847460
|
| [3] |
Turk DC (1999) The role of psychological factors in chronic pain. Acta Anaesthesiol Scand 43: 885-888. doi: 10.1034/j.1399-6576.1999.430904.x
|
| [4] |
Meints SM, Edwards RR (2018) Evaluating psychosocial contributions to chronic pain outcomes. Prog Neuropsychopharmacol Biol Psychiatry 87: 168-182. doi: 10.1016/j.pnpbp.2018.01.017
|
| [5] |
Gatchel RJ, McGeary DD, McGeary CA, et al. (2014) Interdisciplinary chronic pain management: past, present, and future. Am psychol 69: 119-130. doi: 10.1037/a0035514
|
| [6] |
Gatchel RJ (2005) Clinical essentials of pain management American Psychological Association. doi: 10.1037/10856-000
|
| [7] |
Bevers K, Watts L, Kishino ND, et al. (2016) The biopsychosocial model of the assessment, prevention, and treatment of chronic pain. US Neurol 12: 98-104. doi: 10.17925/USN.2016.12.02.98
|
| [8] |
Burke NN, Finn DP, McGuire BE, et al. (2017) Psychological stress in early life as a predisposing factor for the development of chronic pain: clinical and preclinical evidence and neurobiological mechanisms. J Neurosci Res 95: 1257-1270. doi: 10.1002/jnr.23802
|
| [9] |
Sturgeon JA, Zautra AJ (2016) Social pain and physical pain: shared paths to resilience. Pain Manag 6: 63-74. doi: 10.2217/pmt.15.56
|
| [10] |
Gupta A, Mayer EA, Fling C, et al. (2017) Sex-based differences in brain alterations across chronic pain conditions. J Neurosci Res 95: 604-616. doi: 10.1002/jnr.23856
|
| [11] |
Tanasescu R, Cottam WJ, Condon L, et al. (2016) Functional reorganization in chronic pain and neural correlates of pain sensitization: a coordinate based meta-analysis of 266 cutaneous pain fMRI studies. Neurosci Biobehav Rev 68: 120-133. doi: 10.1016/j.neubiorev.2016.04.001
|
| [12] |
Jegindø EME, Vase L, Skewes JC, et al. (2013) Expectations contribute to reduced pain levels during prayer in highly religious participants. J Behav Med 36: 413-426. doi: 10.1007/s10865-012-9438-9
|
| [13] |
Edwards RR, Moric M, Husfeldt B, et al. (2005) Ethnic similarities and differences in the chronic pain experience: a comparison of African American, Hispanic, and white patients. Pain Med 6: 88-98. doi: 10.1111/j.1526-4637.2005.05007.x
|
| [14] |
Kregel J, Meeus M, Malfliet A, et al. (2015) Structural and functional brain abnormalities in chronic low back pain: A systematic review. Semin Arthritis Rheum 45: 229-237. doi: 10.1016/j.semarthrit.2015.05.002
|
| [15] |
Seminowicz DA, Moayedi M (2017) The dorsolateral prefrontal cortex in acute and chronic pain. J Pain 18: 1027-1035. doi: 10.1016/j.jpain.2017.03.008
|
| [16] |
May A (2008) Chronic pain may change the structure of the brain. Pain 137: 7-15. doi: 10.1016/j.pain.2008.02.034
|
| [17] |
Rodriguez-Raecke R, Niemeier A, Ihle K, et al. (2013) Structural brain changes in chronic pain reflect probably neither damage nor atrophy. PloS One 8: e54475. doi: 10.1371/journal.pone.0054475
|
| [18] |
Peirs C, Seal RP (2016) Neural circuits for pain: recent advances and current views. Science 354: 578-584. doi: 10.1126/science.aaf8933
|
| [19] | Hendry SH, Hsiao S (2012) The somatosensory system. Fundamental neuroscience New York: Elsevier, 531-551. |
| [20] |
Kuner R, Flor H (2017) Structural plasticity and reorganisation in chronic pain. Nat Rev Neurosci 18: 20. doi: 10.1038/nrn.2016.162
|
| [21] |
Cai YQ, Wang W, Paulucci-Holthauzen A, et al. (2018) Brain circuits mediating opposing effects on emotion and pain. J Neurosci 38: 6340-6349. doi: 10.1523/JNEUROSCI.2780-17.2018
|
| [22] |
Schweinhardt P, Bushnell MC (2010) Pain imaging in health and disease—how far have we come? J Clin Invest 120: 3788-3797. doi: 10.1172/JCI43498
|
| [23] |
Demirtaş M, Burt JB, Helmer M, et al. (2019) Hierarchical heterogeneity across human cortex shapes large-scale neural dynamics. Neuron 101: 1181-1194. doi: 10.1016/j.neuron.2019.01.017
|
| [24] | Woo CW, Koban L, Kross E, et al. (2014) Separate neural representations for physical pain and social rejection. Nat Commun 5: 1-12. |
| [25] |
Wiech K (2016) Deconstructing the sensation of pain: The influence of cognitive processes on pain perception. Science 354: 584-587. doi: 10.1126/science.aaf8934
|
| [26] |
Reddan MC, Wager TD (2018) Modeling pain using fMRI: from regions to biomarkers. Neurosci Bull 34: 208-215. doi: 10.1007/s12264-017-0150-1
|
| [27] |
Tatu K, Costa T, Nani A, et al. (2018) How do morphological alterations caused by chronic pain distribute across the brain? A meta-analytic co-alteration study. Neuroimage Clin 18: 15-30. doi: 10.1016/j.nicl.2017.12.029
|
| [28] |
Woo CW, Schmidt L, Krishnan A, et al. (2017) Quantifying cerebral contributions to pain beyond nociception. Nat Commun 8: 1-14. doi: 10.1038/s41467-016-0009-6
|
| [29] |
Woo CW, Roy M, Buhle JT, et al. (2015) Distinct brain systems mediate the effects of nociceptive input and self-regulation on pain. PLoS Biol 13: e1002036. doi: 10.1371/journal.pbio.1002036
|
| [30] |
Alshelh Z, Marciszewski KK, Akhter RD, et al. (2018) Disruption of default mode network dynamics in acute and chronic pain states. Neuroimage Clin 17: 222-231. doi: 10.1016/j.nicl.2017.10.019
|
| [31] |
Zhang Y, Zhou W, Wang S, et al. (2019) The roles of subdivisions of human insula in emotion perception and auditory processing. Cereb Cortex 29: 517-528. doi: 10.1093/cercor/bhx334
|
| [32] |
Chiang MC, Bowen A, Schier LA, et al. (2019) Parabrachial Complex: A Hub for pain and aversion. J Neurosci 39: 8225-8230. doi: 10.1523/JNEUROSCI.1162-19.2019
|
| [33] |
Da Silva JT, Seminowicz DA (2019) Neuroimaging of pain in animal models: A review of recent literature. Pain Rep 4: e732. doi: 10.1097/PR9.0000000000000732
|
| [34] | Luria AR (1966) Higher cortical functions in man New York: Basic Books. |
| [35] | Moss RA (2006) Of bits and logic: Cortical columns in learning and memory. J Mind Behav 27: 215-246. |
| [36] |
Moss RA (2016) A theory on the singular function of the hippocampus: Facilitating the binding of new circuits of cortical columns. AIMS Neurosci 3: 264-305. doi: 10.3934/Neuroscience.2016.3.264
|
| [37] | Moss RA (2013a) Psychotherapy and the brain: The dimensional systems model and clinical biopsychology. J Mind Behav 34: 63-89. |
| [38] | Calvin WH (1995) Cortical columns, modules, and Hebbian cell assemblies. Handbook of brain theory and neural networks Cambridge, Massachusetts: MIT Press, 269-275. |
| [39] | Moss RA, Hunter BP, Shah D, et al. (2012) A theory of hemispheric specialization based on cortical columns. J Mind Behav 33: 141-171. |
| [40] |
Horton JC, Adams DL (2005) The cortical column: A structure without a function. Philos Trans R Soc B Biol Sci 360: 837-862. doi: 10.1098/rstb.2005.1623
|
| [41] |
Moss RA, Moss J (2014) The role of cortical columns in explaining gamma-band synchronization and NMDA receptors in cognitive functions. AIMS Neurosci 1: 65-88. doi: 10.3934/Neuroscience.2014.1.65
|
| [42] |
Cheng K, Waggoner RA, Tanaka K (2001) Human ocular dominance columns as revealed by high-field functional magnetic resonance imaging. Neuron 32: 359-374. doi: 10.1016/S0896-6273(01)00477-9
|
| [43] |
Yacoub E, Harel N, Uğurbil K (2008) High-field fMRI unveils orientation columns in humans. Proc Natl Acad Sci 105: 10607-10612. doi: 10.1073/pnas.0804110105
|
| [44] |
Nasr S, Polimeni JR, Tootell RB (2016) Interdigitated color-and disparity-selective columns within human visual cortical areas V2 and V3. J Neurosci 36: 1841-1857. doi: 10.1523/JNEUROSCI.3518-15.2016
|
| [45] |
Goncalves NR, Ban H, Sánchez-Panchuelo RM, et al. (2015) 7 tesla FMRI reveals systematic functional organization for binocular disparity in dorsal visual cortex. J Neurosci 35: 3056-3072. doi: 10.1523/JNEUROSCI.3047-14.2015
|
| [46] |
Zimmermann J, Goebel R, De Martino F, et al. (2011) Mapping the organization of axis of motion selective features in human area MT using high-field fMRI. PloS One 6: e28716. doi: 10.1371/journal.pone.0028716
|
| [47] |
Margalit E, Jamison KW, Weiner KS, et al. (2020) Ultra-high-resolution fMRI of human ventral temporal cortex reveals differential representation of categories and domains. J Neurosci 40: 3008-3024. doi: 10.1523/JNEUROSCI.2106-19.2020
|
| [48] |
Schneider M, Kemper VG, Emmerling TC, et al. (2019) Columnar clusters in the human motion complex reflect consciously perceived motion axis. Proc Natl Acad Sci 116: 5096-5101. doi: 10.1073/pnas.1814504116
|
| [49] |
Crick F, Koch C (2003) A framework for consciousness. Nat Neurosci 6: 119-126. doi: 10.1038/nn0203-119
|
| [50] |
Gazzaniga MS (2002) Consciousness. Encyclopedia of the human brain New York: Academic Press, 31-35. doi: 10.1016/B0-12-227210-2/00106-0
|
| [51] | Moss RA (1992) Emotional restructuring: A structured psychotherapeutic approach for depression and anxiety Greenville, SC: St. Francis Hospital. |
| [52] | Moss RA (2001) Clinical biopsychology in theory and practice Greenville, South Carolina: Center for Emotional Restructuring. |
| [53] |
Moss RA (2007) Negative emotional memories in clinical treatment: Theoretical considerations. J Psychother Integration 17: 209-224. doi: 10.1037/1053-0479.17.2.209
|
| [54] | Moss RA (2015) Psychotherapy integration from a brain-based perspective: Clinical biopsychology. Continuing education course offered through Health Forum Online.Available from: www.healthforumonline.com. |
| [55] |
Moss RA (2013b) Givers and takers: Clinical biopsychological perspectives on relationship behavior patterns. Int J Neuropsychother 1: 31-46. doi: 10.12744/ijnpt.2013.0031-0046
|
| [56] |
Moss RA (2013c) A clinical biopsychological theory of loss-related depression. Int J Neuropsychother 1: 56-65. doi: 10.12744/ijnpt.2013.0056-0065
|
| [57] |
Jung J, Cloutman LL, Binney RJ, et al. (2017) The structural connectivity of higher order association cortices reflects human functional brain networks. Cortex 97: 221-239. doi: 10.1016/j.cortex.2016.08.011
|
| [58] |
Jung J, Visser M, Binney RJ, et al. (2018) Establishing the cognitive signature of human brain networks derived from structural and functional connectivity. Brain Struct Funct 223: 4023-4038. doi: 10.1007/s00429-018-1734-x
|
| [59] | Purves D, Augustine GJ, Fitzpatrick D, et al. (2012) Neuroscience Massachusetts: Sinauer Associates, Inc. |
| [60] |
Hélie S, Ell SW, Ashby FG (2015) Learning robust cortico-cortical associations with the basal ganglia: An integrative review. Cortex 64: 123-135. doi: 10.1016/j.cortex.2014.10.011
|
| [61] |
Cacciola A, Milardi D, Bertino S, et al. (2019) Structural connectivity-based topography of the human globus pallidus: Implications for therapeutic targeting in movement disorders. Mov Disord 34: 987-996. doi: 10.1002/mds.27712
|
| [62] |
Vogelsang DA, D'Esposito M (2018) Is there evidence for a rostral-caudal gradient in fronto-striatal loops and what role does dopamine play? Front Neurosci 12: 242. doi: 10.3389/fnins.2018.00242
|
| [63] |
Pauli WM, O'Reilly RC, Yarkoni T, et al. (2016) Regional specialization within the human striatum for diverse psychological functions. Proc Natl Acad Sci 113: 1907-1912. doi: 10.1073/pnas.1507610113
|
| [64] |
Haber SN (2003) The primate basal ganglia: parallel and integrative networks. J Chem Neuroanat 26: 317-330. doi: 10.1016/j.jchemneu.2003.10.003
|
| [65] |
Ghazizadeh A, Hong S, Hikosaka O (2018) Prefrontal cortex represents long-term memory of object values for months. Curr Biol 28: 2206-2217. doi: 10.1016/j.cub.2018.05.017
|
| [66] |
Ceceli AO, Tricomi E (2018) Habits and goals: A motivational perspective on action control. Curr Opin Behav Sci 20: 110-116. doi: 10.1016/j.cobeha.2017.12.005
|
| [67] |
Soares JM, Sampaio A, Ferreira LM, et al. (2012) Stress-induced changes in human decision-making are reversible. Transl Psychiatry 2: e131. doi: 10.1038/tp.2012.59
|
| [68] | LeDoux JL, Phelps EA (2010) Emotional networks in the brain. Handbook of emotions New York: Guilford Press, 159-179. |
| [69] |
Milardi D, Arrigo A, Anastasi G, et al. (2016) Extensive direct subcortical cerebellum-basal ganglia connections in human brain as revealed by constrained spherical deconvolution tractography. Front Neuroanat 10: 29. doi: 10.3389/fnana.2016.00029
|
| [70] |
Aron AR, Herz DM, Brown P, et al. (2016) Frontosubthalamic circuits for control of action and cognition. J Neurosci 36: 11489-11495. doi: 10.1523/JNEUROSCI.2348-16.2016
|
| [71] |
Cacciola A, Calamuneri A, Milardi D, et al. (2017) A connectomic analysis of the human basal ganglia network. Front Neuroanat 11: 85. doi: 10.3389/fnana.2017.00085
|
| [72] |
Snow WM, Stoesz BM, Anderson JE (2014) The cerebellum in emotional processing: evidence from human and non-human animals. Aims Neurosci 1: 96-119. doi: 10.3934/Neuroscience.2014.1.96
|
| [73] |
Balsters JH, Laird AR, Fox PT, et al. (2014) Bridging the gap between functional and anatomical features of cortico-cerebellar circuits using meta-analytic connectivity modeling. Hum Brain Mapp 35: 3152-3169. doi: 10.1002/hbm.22392
|
| [74] |
Nakamura Y, Kato H, Nojiri K, et al. (2014) Significant differences of brain blood flow in patients with chronic low back pain and acute low back pain detected by brain SPECT. J Orthop Sci 19: 384-389. doi: 10.1007/s00776-014-0534-2
|
| [75] |
Schmahmann JD, Sherman JC (1997) Cerebellar cognitive affective syndrome. Int Rev Neurobiol 41: 433-440. doi: 10.1016/S0074-7742(08)60363-3
|
| [76] |
Stoodley CJ, Valera EM, Schmahmann JD (2012) Functional topography of the cerebellum for motor and cognitive tasks: An fMRI study. Neuroimage 59: 1560-1570. doi: 10.1016/j.neuroimage.2011.08.065
|
| [77] | Bernard JA, Seidler RD, Hassevoort KM, et al. (2012) Resting state cortico-cerebellar functional connectivity networks: A comparison of anatomical and self-organizing map approaches. Fron Neuroanat 6: 31. |
| [78] |
Schmahmann JD (2004) Disorders of the cerebellum: ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J Neuropsychiatry Clin Neurosci 16: 367-378. doi: 10.1176/jnp.16.3.367
|
| [79] |
Guell X, Gabrieli JD, Schmahmann JD (2018) Embodied cognition and the cerebellum: Perspectives from the dysmetria of thought and the universal cerebellar transform theories. Cortex 100: 140-148. doi: 10.1016/j.cortex.2017.07.005
|
| [80] |
Milardi D, Gaeta M, Marino S, et al. (2015) Basal ganglia network by constrained spherical deconvolution: a possible cortico-pallidal pathway? Mov Disord 30: 342-349. doi: 10.1002/mds.25995
|
| [81] |
Watson P, van Wingen G, de Wit S (2018) Conflicted between goal-directed and habitual control, an fMRI investigation. eNeuro 5. doi: 10.1523/ENEURO.0240-18.2018
|
| [82] |
Joyce MKP, Barbas H (2018) Cortical connections position primate area 25 as a keystone for interoception, emotion, and memory. J Neurosci 38: 1677-1698. doi: 10.1523/JNEUROSCI.2363-17.2017
|
| [83] |
Ghashghaei HT, Hilgetag CC, Barbas H (2007) Sequence of information processing for emotions based on the anatomic dialogue between prefrontal cortex and amygdala. Neuroimage 34: 905-923. doi: 10.1016/j.neuroimage.2006.09.046
|
| [84] |
Ray RD, Zald DH (2012) Anatomical insights into the interaction of emotion and cognition in the prefrontal cortex. Neurosci Biobehav Rev 36: 479-501. doi: 10.1016/j.neubiorev.2011.08.005
|
| [85] |
Ji RR, Chamessian A, Zhang YQ (2016) Pain regulation by non-neuronal cells and inflammation. Science 354: 572-577. doi: 10.1126/science.aaf8924
|
| [86] |
Kuner R, Flor H (2017) Structural plasticity and reorganisation in chronic pain. Nat Rev Neurosci 18: 20. doi: 10.1038/nrn.2016.162
|
| [87] |
Sommer C (2016) Exploring pain pathophysiology in patients. Science 354: 588-592. doi: 10.1126/science.aaf8935
|
| [88] |
Chang PC, Centeno MV, Procissi D, et al. (2017) Brain activity for tactile allodynia: a longitudinal awake rat functional magnetic resonance imaging study tracking emergence of neuropathic pain. Pain 158: 488-497. doi: 10.1097/j.pain.0000000000000788
|
| [89] |
Geha PY, Baliki MN, Wang X, et al. (2008) Brain dynamics for perception of tactile allodynia (touch-induced pain) in postherpetic neuralgia. Pain 138: 641-656. doi: 10.1016/j.pain.2008.02.021
|
| [90] |
Lanz S, Seifert F, Maihöfner C (2011) Brain activity associated with pain, hyperalgesia and allodynia: an ALE meta-analysis. J Neur Transmis 118: 1139-1154. doi: 10.1007/s00702-011-0606-9
|
| [91] |
Pavuluri M, May A (2015) I feel, therefore, I am: the insula and its role in human emotion, cognition and the sensory-motor system. AIMS Neurosci 2: 18-27. doi: 10.3934/Neuroscience.2015.1.18
|
| [92] |
Cloutman LL, Binney RJ, Drakesmith M, et al. (2012) The variation of function across the human insula mirrors its patterns of structural connectivity: evidence from in vivo probabilistic tractography. Neuroimage 59: 3514-3521. doi: 10.1016/j.neuroimage.2011.11.016
|
| [93] |
Olausson H, Wessberg J, McGlone F, et al. (2010) The neurophysiology of unmyelinated tactile afferents. Neurosci Biobehav Rev 34: 185-191. doi: 10.1016/j.neubiorev.2008.09.011
|
| [94] |
Björnsdotter M, Olausson H (2011) Vicarious responses to social touch in posterior insular cortex are tuned to pleasant caressing speeds. J Neurosci 31: 9554-9562. doi: 10.1523/JNEUROSCI.0397-11.2011
|
| [95] |
Davidovic M, Starck G, Olausson H (2019) Processing of affective and emotionally neutral tactile stimuli in the insular cortex. Dev Cogn Neurosci 35: 94-103. doi: 10.1016/j.dcn.2017.12.006
|
| [96] | Craig AD (2010) Interoception and emotion: A neuroanatomical perspective. Handbook of emotions New York: Guilford Press, 272-292. |
| [97] |
Frot M, Magnin M, Mauguière F, et al. (2006) Human SII and posterior insula differently encode thermal laser stimuli. Cereb Cortex 17: 610-620. doi: 10.1093/cercor/bhk007
|
| [98] |
Baliki MN, Geha PY, Apkarian AV (2009) Parsing pain perception between nociceptive representation and magnitude estimation. J Neurophysiol 101: 875-887. doi: 10.1152/jn.91100.2008
|
| [99] |
Zhang Y, Zhou W, Wang S, et al. (2019) The roles of subdivisions of human insula in emotion perception and auditory processing. Cereb Cortex 29: 517-528. doi: 10.1093/cercor/bhx334
|
| [100] |
Jackson RL, Bajada CJ, Lambon Ralph MA, et al. (2019) The graded change in connectivity across the ventromedial prefrontal cortex reveals distinct subregions. Cereb Cortex 30: 165-180. doi: 10.1093/cercor/bhz079
|
| [101] |
Bastuji H, Frot M, Perchet C, et al. (2018) Convergence of sensory and limbic noxious input into the anterior insula and the emergence of pain from nociception. Sci Rep 8: 1-9. doi: 10.1038/s41598-018-31781-z
|
| [102] |
Nomi JS, Farrant K, Damaraju E, et al. (2016) Dynamic functional network connectivity reveals unique and overlapping profiles of insula subdivisions. Hum Brain Mapp 37: 1770-1787. doi: 10.1002/hbm.23135
|
| [103] |
Tatu K, Costa T, Nani A, et al. (2018) How do morphological alterations caused by chronic pain distribute across the brain? A meta-analytic co-alteration study. Neuroimage Clin 18: 15-30. doi: 10.1016/j.nicl.2017.12.029
|
| [104] |
Vogt BA (2005) Pain and emotion interactions in subregions of the cingulate gyrus. Nat Rev Neurosci 6: 533-544. doi: 10.1038/nrn1704
|
| [105] |
Beckmann M, Johansen-Berg H, Rushworth MF (2009) Connectivity-based parcellation of human cingulate cortex and its relation to functional specialization. J Neurosci 29: 1175-1190. doi: 10.1523/JNEUROSCI.3328-08.2009
|
| [106] |
Vogt BA (2016) Midcingulate cortex: structure, connections, homologies, functions and diseases. J Chem Neuroanat 74: 28-46. doi: 10.1016/j.jchemneu.2016.01.010
|
| [107] |
Leech R, Sharp DJ (2014) The role of the posterior cingulate cortex in cognition and disease. Brain 137: 12-32. doi: 10.1093/brain/awt162
|
| [108] |
Fischer LF, Soto-Albors RM, Buck F, et al. (2020) Representation of visual landmarks in retrosplenial cortex. Elife 9: e51458. doi: 10.7554/eLife.51458
|
| [109] |
Tang W, Jbabdi S, Zhu Z, et al. (2019) A connectional hub in the rostral anterior cingulate cortex links areas of emotion and cognitive control. Elife 8: e43761. doi: 10.7554/eLife.43761
|
| [110] |
de la Vega A, Chang LJ, Banich MT, et al. (2016) Large-scale meta-analysis of human medial frontal cortex reveals tripartite functional organization. J Neurosci 36: 6553-6562. doi: 10.1523/JNEUROSCI.4402-15.2016
|
| [111] |
Li YS, Nassar MR, Kable JW, et al. (2019) Individual neurons in the cingulate cortex encode action monitoring, not selection, during adaptive decision-making. J Neurosci 39: 6668-6683. doi: 10.1523/JNEUROSCI.0159-19.2019
|
| [112] |
Shenhav A, Cohen JD, Botvinick MM (2016) Dorsal anterior cingulate cortex and the value of control. Nat Neurosci 19: 1286-1291. doi: 10.1038/nn.4384
|
| [113] |
Kolling N, Behrens TEJ, Wittmann MK, et al. (2016) Multiple signals in anterior cingulate cortex. Curr Opin Neurobiol 37: 36-43. doi: 10.1016/j.conb.2015.12.007
|
| [114] |
Holroyd CB, Yeung N (2012) Motivation of extended behaviors by anterior cingulate cortex. Trends Cogn Sci 16: 122-128. doi: 10.1016/j.tics.2011.12.008
|
| [115] |
Brockett AT, Tennyson SS, deBettencourt CA, et al. (2020) Anterior cingulate cortex is necessary for adaptation of action plans. Proc Natl Acad Sci 117: 6196-6204. doi: 10.1073/pnas.1919303117
|
| [116] |
Tomasi D, Volkow ND (2011) Association between functional connectivity hubs and brain networks. Cereb Cortex 21: 2003-2013. doi: 10.1093/cercor/bhq268
|
| [117] |
Margulies DS, Vincent JL, Kelly C, et al. (2009) Precuneus shares intrinsic functional architecture in humans and monkeys. Proc Natl Acad Sci 106: 20069-20074. doi: 10.1073/pnas.0905314106
|
| [118] |
Cavanna AE, Trimble MR (2006) The precuneus: a review of its functional anatomy and behavioural correlates. Brain 129: 564-583. doi: 10.1093/brain/awl004
|
| [119] |
Utevsky AV, Smith DV, Huettel SA (2014) Precuneus is a functional core of the default-mode network. J Neurosci 34: 932-940. doi: 10.1523/JNEUROSCI.4227-13.2014
|
| [120] |
Foudil SA, Kwok SC, Macaluso E (2020) Context-dependent coding of temporal distance between cinematic events in the human precuneus. J Neurosci 40: 2129-2138. doi: 10.1523/JNEUROSCI.2296-19.2020
|
| [121] |
Lamm C, Decety J, Singer T (2011) Meta-analytic evidence for common and distinct neural networks associated with directly experienced pain and empathy for pain. Neuroimage 54: 2492-2502. doi: 10.1016/j.neuroimage.2010.10.014
|
| [122] |
Badre D, D'Esposito M (2009) Is the rostro-caudal axis of the frontal lobe hierarchical? Nat Rev Neurosci 10: 659-669. doi: 10.1038/nrn2667
|
| [123] |
Petrides M, Pandya DN (1984) Projections to the frontal cortex from the posterior parietal region in the rhesus monkey. J Comp Neurol 228: 105-116. doi: 10.1002/cne.902280110
|
| [124] | Riley MR, Qi XL, Constantinidis C (2017) Functional specialization of areas along the anterior–posterior axis of the primate prefrontal cortex. Cereb Cortex 27: 3683-3697. |
| [125] |
Ong WY, Stohler CS, Herr DR (2019) Role of the prefrontal cortex in pain processing. Mol Neurobiol 56: 1137-1166. doi: 10.1007/s12035-018-1130-9
|
| [126] |
Duncan J (2010) The multiple-demand (MD) system of the primate brain: Mental programs for intelligent behaviour. Trends Cogn Sci 14: 172-179. doi: 10.1016/j.tics.2010.01.004
|
| [127] |
Hempel A, Giesel FL, Garcia Caraballo NM, et al. (2004) Plasticity of cortical activation related to working memory during training. Am J Psychiatry 161: 745-747. doi: 10.1176/appi.ajp.161.4.745
|
| [128] |
Sevel LS, Letzen JE, Staud R, et al. (2016) Interhemispheric dorsolateral prefrontal cortex connectivity is associated with individual differences in pain sensitivity in healthy controls. Brain Connect 6: 357-364. doi: 10.1089/brain.2015.0405
|
| [129] |
Korb FM, Jiang J, King JA, et al. (2017) Hierarchically organized medial frontal cortex-basal ganglia loops selectively control task-and response-selection. J Neurosci 37: 7893-7905. doi: 10.1523/JNEUROSCI.3289-16.2017
|
| [130] |
Kahnt T, Chang LJ, Park SQ, et al. (2012) Connectivity-based parcellation of the human orbitofrontal cortex. J Neurosci 32: 6240-6250. doi: 10.1523/JNEUROSCI.0257-12.2012
|
| [131] |
Costa VD, Averbeck BB (2020) Primate orbitofrontal cortex codes information relevant for managing explore–exploit tradeoffs. J Neurosci 40: 2553-2561. doi: 10.1523/JNEUROSCI.2355-19.2020
|
| [132] |
Samara Z, Evers EA, Goulas A, et al. (2017) Human orbital and anterior medial prefrontal cortex: Intrinsic connectivity parcellation and functional organization. Brain Struct Funct 222: 2941-2960. doi: 10.1007/s00429-017-1378-2
|
| [133] |
Phillips ML, Ladouceur CD, Drevets WC (2008) A neural model of voluntary and automatic emotion regulation: implications for understanding the pathophysiology and neurodevelopment of bipolar disorder. Mol Psychiatry 13: 833-857. doi: 10.1038/mp.2008.65
|
| [134] |
Vachon-Presseau E, Tétreault P, Petre B, et al. (2016) Corticolimbic anatomical characteristics predetermine risk for chronic pain. Brain 139: 1958-1970. doi: 10.1093/brain/aww100
|
| [135] |
Hibar DP, Stein JL, Renteria ME, et al. (2015) Common genetic variants influence human subcortical brain structures. Nature 520: 224-229. doi: 10.1038/nature14101
|
| [136] |
Gilbertson MW, Shenton ME, Ciszewski A, et al. (2002) Smaller hippocampal volume predicts pathologic vulnerability to psychological trauma. Nat Neurosci 5: 1242-1247. doi: 10.1038/nn958
|
| [137] |
Oshri A, Gray JC, Owens MM, et al. (2019) Adverse childhood experiences and amygdalar reduction: High-resolution segmentation reveals associations with subnuclei and psychiatric outcomes. Child Maltreat 24: 400-410. doi: 10.1177/1077559519839491
|
| [138] |
Johnson FK, Delpech JC, Thompson GJ, et al. (2018) Amygdala hyper-connectivity in a mouse model of unpredictable early life stress. Transl Psychiatry 8: 1-14. doi: 10.1038/s41398-018-0092-z
|
| [139] |
Hashmi JA, Baliki MN, Huang L, et al. (2013) Shape shifting pain: Chronification of back pain shifts brain representation from nociceptive to emotional circuits. Brain 136: 2751-2768. doi: 10.1093/brain/awt211
|
| [140] |
Diano M, Tamietto M, Celeghin A, et al. (2017) Dynamic changes in amygdala psychophysiological connectivity reveal distinct neural networks for facial expressions of basic emotions. Sci Rep 7: 45260. doi: 10.1038/srep45260
|
| [141] |
Sander D, Grafman J, Zalla T (2003) The human amygdala: An evolved system for relevance detection. Rev Neurosci 14: 303-316. doi: 10.1515/REVNEURO.2003.14.4.303
|
| [142] |
Kerestes R, Chase HW, Phillips ML, et al. (2017) Multimodal evaluation of the amygdala's functional connectivity. Neuroimage 148: 219-229. doi: 10.1016/j.neuroimage.2016.12.023
|
| [143] |
Balleine BW, Killcross S (2006) Parallel incentive processing: An integrated view of amygdala function. Trends Neurosci 29: 272-279. doi: 10.1016/j.tins.2006.03.002
|
| [144] |
Roy AK, Shehzad Z, Margulies DS, et al. (2009) Functional connectivity of the human amygdala using resting state fMRI. Neuroimage 45: 614-626. doi: 10.1016/j.neuroimage.2008.11.030
|
| [145] |
Bzdok D, Laird AR, Zilles K, et al. (2013) An investigation of the structural, connectional, and functional subspecialization in the human amygdala. Hum Brain Mapp 34: 3247-3266. doi: 10.1002/hbm.22138
|
| [146] |
Gorka AX, Torrisi S, Shackman AJ, et al. (2018) Intrinsic functional connectivity of the central nucleus of the amygdala and bed nucleus of the stria terminalis. Neuroimage 168: 392-402. doi: 10.1016/j.neuroimage.2017.03.007
|
| [147] |
Bandler R, Keay KA, Floyd N, et al. (2000) Central circuits mediating patterned autonomic activity during active vs. passive emotional coping. Brain Res Bull 53: 95-104. doi: 10.1016/S0361-9230(00)00313-0
|
| [148] |
Oler JA, Tromp DP, Fox AS, et al. (2017) Connectivity between the central nucleus of the amygdala and the bed nucleus of the stria terminalis in the non-human primate: neuronal tract tracing and developmental neuroimaging studies. Brain Struct Funct 222: 21-39. doi: 10.1007/s00429-016-1198-9
|
| [149] |
Folloni D, Sallet J, Khrapitchev AA, et al. (2019) Dichotomous organization of amygdala/temporal-prefrontal bundles in both humans and monkeys. Elife 8: e47175. doi: 10.7554/eLife.47175
|
| [150] |
Schmidt-Wilcke T, Luerding R, Weigand T, et al. (2007) Striatal grey matter increase in patients suffering from fibromyalgia–a voxel-based morphometry study. Pain 132: S109-S116. doi: 10.1016/j.pain.2007.05.010
|
| [151] |
Diano M, D'Agata F, Cauda F, et al. (2016) Cerebellar clustering and functional connectivity during pain processing. Cerebellum 15: 343-356. doi: 10.1007/s12311-015-0706-4
|
| [152] |
Coombes SA, Misra G (2016) Pain and motor processing in the human cerebellum. Pain 157: 117-127. doi: 10.1097/j.pain.0000000000000337
|
| [153] |
Moulton EA, Elman I, Pendse G, et al. (2011) Aversion-related circuitry in the cerebellum: responses to noxious heat and unpleasant images. J Neurosci 31: 3795-3804. doi: 10.1523/JNEUROSCI.6709-10.2011
|
| [154] |
Kim H, Kim J, Loggia ML, et al. (2015) Fibromyalgia is characterized by altered frontal and cerebellar structural covariance brain networks. Neuroimage Clin 7: 667-677. doi: 10.1016/j.nicl.2015.02.022
|
| [155] |
Tsutsumi S, Hidaka N, Isomura Y, et al. (2019) Modular organization of cerebellar climbing fiber inputs during goal-directed behavior. Elife 8: e47021. doi: 10.7554/eLife.47021
|
| [156] |
Larry N, Yarkoni M, Lixenberg A, et al. (2019) Cerebellar climbing fibers encode expected reward size. Elife 8: e46870. doi: 10.7554/eLife.46870
|
| [157] |
Heffley W, Hull C (2019) Classical conditioning drives learned reward prediction signals in climbing fibers across the lateral cerebellum. Elife 8: e46764. doi: 10.7554/eLife.46764
|
| [158] |
Watson TC, Obiang P, Torres-Herraez A, et al. (2019) Anatomical and physiological foundations of cerebello-hippocampal interaction. Elife 8: e41896. doi: 10.7554/eLife.41896
|
| [159] |
Apkarian AV, Mutso AA, Centeno MV, et al. (2016) Role of adult hippocampal neurogenesis in persistent pain. Pain 157: 418-428. doi: 10.1097/j.pain.0000000000000332
|
| [160] |
Mutso AA, Radzicki D, Baliki MN, et al. (2012) Abnormalities in hippocampal functioning with persistent pain. J Neurosci 32: 5747-5756. doi: 10.1523/JNEUROSCI.0587-12.2012
|
| [161] |
Mutso AA, Petre B, Huang L, et al. (2014) Reorganization of hippocampal functional connectivity with transition to chronic back pain. J Neurophysiol 111: 1065-1076. doi: 10.1152/jn.00611.2013
|
| [162] |
Borsook D, Linnman C, Faria V, et al. (2016) Reward deficiency and anti-reward in pain chronification. Neurosci Biobehav Rev 68: 282-297. doi: 10.1016/j.neubiorev.2016.05.033
|
| [163] |
DosSantos MF, Moura BDS, DaSilva AF (2017) Reward circuitry plasticity in pain perception and modulation. Front Pharmacol 8: 790. doi: 10.3389/fphar.2017.00790
|
| [164] |
Harris HN, Peng YB (2020) Evidence and explanation for the involvement of the nucleus accumbens in pain processing. Neuro Regen Res 15: 597-605. doi: 10.4103/1673-5374.266909
|
| [165] |
Yu R, Gollub RL, Spaeth R, et al. (2014) Disrupted functional connectivity of the periaqueductal gray in chronic low back pain. Neuroimage Clin 6: 100-108. doi: 10.1016/j.nicl.2014.08.019
|
| [166] | Li Y, Wang Y, Xuan C, et al. (2017) Role of the lateral habenula in pain-associated depression. Front Behav Neurosci 11: 31. |
| [167] |
Shelton L, Becerra L, Borsook D (2012) Unmasking the mysteries of the habenula in pain and analgesia. Prog Neurobiol 96: 208-219. doi: 10.1016/j.pneurobio.2012.01.004
|
| [168] |
Shelton L, Pendse G, Maleki N, et al. (2012) Mapping pain activation and connectivity of the human habenula. J Neurophysiol 107: 2633-2648. doi: 10.1152/jn.00012.2012
|
| [169] |
Kaschube M, Schnabel M, Wolf F, et al. (2009) Interareal coordination of columnar architectures during visual cortical development. Proc Natl Acad Sci 106: 17205-17210. doi: 10.1073/pnas.0901615106
|
| [170] | Stermensky II G, Moss RA (2017) Cognitive symptoms and effects of stress. Stress in the modern world: Understanding science and society Santa Barbara, CA: ABC-CLIO. |
| [171] |
Ecker B (2015) Memory reconsolidation understood and misunderstood. Int J Neuropsychother 3: 2-46. doi: 10.12744/ijnpt.2015.0002-0046
|
| [172] |
Bossert JM, Stern AL, Theberge FRM, et al. (2012) Role of projections from ventral medial prefrontal cortex to nucleus accumbens shell in context-induced reinstatement of heroin seeking. J Neurosci 32: 4982-4991. doi: 10.1523/JNEUROSCI.0005-12.2012
|
| [173] |
Hecht D (2010) Depression and the hyperactive right-hemisphere. Neurosci Res 68: 77-87. doi: 10.1016/j.neures.2010.06.013
|
| [174] |
Rogers CR (1957) The necessary and sufficient conditions of therapeutic personality change. J Consult Psychol 21: 95-103. doi: 10.1037/h0045357
|
| [175] |
Theobald EJ, Hill MJ, Tran E, et al. (2020) Active learning narrows achievement gaps for underrepresented students in undergraduate science, technology, engineering, and math. Proc Natl Acad Sci 117: 6476-6483. doi: 10.1073/pnas.1916903117
|
| [176] |
Hauner KK, Mineka S, Voss JL, et al. (2012) Exposure therapy triggers lasting reorganization of neural fear processing. Proc Natl Acad Sci 109: 9203-9208. doi: 10.1073/pnas.1205242109
|
| [177] |
Cacioppo JT, Berntson GG (1992) Social psychological contributions to the decade of the brain: Doctrine of multilevel analysis. Am Psychol 47: 1019-1028. doi: 10.1037/0003-066X.47.8.1019
|
| [178] |
Adolphs R (2010) Conceptual challenges and directions for social neuroscience. Neuron 65: 752-767. doi: 10.1016/j.neuron.2010.03.006
|
| [179] |
Sliwa J, Freiwald WA (2017) A dedicated network for social interaction processing in the primate brain. Science 356: 745-749. doi: 10.1126/science.aam6383
|
| [180] |
Adolphs R (2001) The neurobiology of social cognition. Curr Opin Neurobiol 11: 231-239. doi: 10.1016/S0959-4388(00)00202-6
|
| [181] | Bates JE, Goodnight JA, Fite JE (2010) Temperament and emotion. Handbook of emotions New York: Guilford Press, 485-496. |
| [182] | Panksepp J (2010) The affective brain and core consciousness: How does neural activity generate emotional feelings? Handbook of emotions New York: Guilford Press, 47-67. |
| [183] |
DeYoung CG, Gray JR (2009) Personality neuroscience: Explaining individual differences in affect, behavior, and cognition. The Cambridge handbook of personality psychology New York: Cambridge University Press, 323-346. doi: 10.1017/CBO9780511596544.023
|
| [184] | Costa PT, McCrae RR (1992) NEO PI-R professional manual Odessa, FL: Psychological Assessment Resources. |
| [185] | John OP, Srivastava S (1999) The Big Five trait taxonomy: History, measurement, and theoretical perspectives. Handbook of personality: Theory and research New York: Guilford, 102-138. |
| [186] |
Jang KL, Livesley WJ, Ando J, et al. (2006) Behavioral genetics of the higher-order factors of the Big Five. Pers Individ Differ 41: 261-272. doi: 10.1016/j.paid.2005.11.033
|
| [187] |
DeYoung CG (2013) The neuromodulator of exploration: A unifying theory of the role of dopamine in personality. Front Hum Neurosci 7: 762. doi: 10.3389/fnhum.2013.00762
|
| [188] |
Wright AG, Creswell KG, Flory JD, et al. (2019) Neurobiological functioning and the personality-trait hierarchy: Central serotonergic responsivity and the stability metatrait. Psychol Sci 30: 1413-1423. doi: 10.1177/0956797619864530
|
| [189] |
DeYoung CG, Hirsh JB, Shane MS, et al. (2010) Testing predictions from personality neuroscience: Brain structure and the Big Five. Psychol Sci 21: 820-828. doi: 10.1177/0956797610370159
|
| [190] |
Wang RW, Chang WL, Chuang SW, et al. (2019) Posterior cingulate cortex can be a regulatory modulator of the default mode network in task-negative state. Sci Rep 9: 1-12. doi: 10.1038/s41598-018-37186-2
|
| [191] |
Fukuda H, Ma N, Suzuki S, et al. (2019) Computing social value conversion in the human brain. J Neurosci 39: 5153-5172. doi: 10.1523/JNEUROSCI.3117-18.2019
|
| [192] |
Seitzman BA, Gratton C, Laumann TO, et al. (2019) Trait-like variants in human functional brain networks. Proc Natl Acad Sci 116: 22851-22861. doi: 10.1073/pnas.1902932116
|
| [193] | Moss RA, Walker GR (2009) For better or for worse: Am I in love with a giver or a taker Greenville, SC: Center for Emotional Restructuring. |
| [194] | Foa EB, Rothbaum BO (1998) Treating the trauma of rape: Cognitive–behavioral therapy for PTSD New York: Guilford. |
| [195] |
Dickerson SS, Kemeny ME (2004) Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychol Bull 130: 355-391. doi: 10.1037/0033-2909.130.3.355
|
| [196] |
Bräscher AK, Becker S, Hoeppli ME, et al. (2016) Different brain circuitries mediating controllable and uncontrollable pain. J Neurosci 36: 5013-5025. doi: 10.1523/JNEUROSCI.1954-15.2016
|
| [197] |
Miller GE, Chen E, Parker KJ (2011) Psychological stress in childhood and susceptibility to the chronic diseases of aging: Moving toward a model of behavioral and biological mechanisms. Psychol Bull 137: 959-997. doi: 10.1037/a0024768
|
| [198] |
Heim C, Wagner D, Maloney E, et al. (2006) Early adverse experience and risk for chronic fatigue syndrome: results from a population-based study. Arch Gen Psychiatry 63: 1258-1266. doi: 10.1001/archpsyc.63.11.1258
|
| [199] |
Bower JE, Crosswell AD, Slavich GM (2014) Childhood adversity and cumulative life stress risk factors for cancer-related fatigue. Clin Psychol Sci 2: 108-115. doi: 10.1177/2167702613496243
|
| [200] | Housley J, Beutler LE (2007) Treating victims of mass disaster and terrorism Cambridge, Massachusetts: Hogrefe. |
| [201] | Greenberg LS (2010) The clinical application of emotion in psychotherapy. Handbook of emotions New York: Guilford, 88-101. |
| [202] | Shapiro F (2017) Eye movement desensitization and reprocessing (EMDR) therapy: Basic principles, protocols, and procedures New York: Guilford Publications. |
| [203] |
Solomon RL (1980) The opponent-process theory of acquired motivation: The costs of pleasure and the benefits of pain. Am Psychol 35: 691-712. doi: 10.1037/0003-066X.35.8.691
|
| [204] |
Hurvich LM, Jameson D (1957) An opponent-process theory of color vision. Psychol Rev 64: 384-404. doi: 10.1037/h0041403
|
| [205] |
Solomon RL, Corbit JD (1973) An opponent-process theory of motivation: II. Cigarette addiction. J Abnorm Psychol 81: 158-171. doi: 10.1037/h0034534
|
| [206] |
Solomon RL, Corbit JD (1974) An opponent-process theory of motivation: I. Temporal dynamics of affect. Psychol Rev 81: 119-145. doi: 10.1037/h0036128
|
| [207] |
Smith DM, Loewenstein G, Jankovic A, et al. (2009) Happily hopeless: Adaptation to a permanent, but not to a temporary, disability. Health Psychol 28: 787-791. doi: 10.1037/a0016624
|
| [208] |
Kirchner T, Lara S (2011) Stress and depression symptoms in patients with multiple sclerosis: The mediating role of loss of social functioning. Acta Neurol Scand 123: 407-413. doi: 10.1111/j.1600-0404.2010.01422.x
|
| [209] |
Nicholson P (1999) Loss, happiness and postpartum depression: the ultimate paradox. Can Psychol 40: 162-178. doi: 10.1037/h0086834
|
| [210] | Lewinsohn PM (1974) Clinical and theoretical aspects of depression. Innovative treatment methods in psychopathology New York: Wiley, 63-120. |
| [211] | Grossberg S, Schmajuk NA (1987) Neural dynamics of attentionally modulated Pavlovian conditioning: Conditioned reinforcement, inhibition, and opponent processing. Psychobiology 15: 195-240. |
| [212] |
Hollerman JR, Schultz W (1998) Dopamine neurons report an error in the temporal prediction of reward during learning. Nat Neurosci 1: 304-309. doi: 10.1038/1124
|
| [213] | Mink JW (2008) The basal ganglia. Fundamental neuroscience New York: Academic Press, 725-750. |
| [214] |
Gundel H, O'Connor MF, Littrell L, et al. (2003) Functional neuroanatomy of grief: An fMRI study. Am J Psychiatry 160: 1946-1953. doi: 10.1176/appi.ajp.160.11.1946
|
| [215] |
O'Connor MF, Wellisch DK, Stanton AL, et al. (2008) Craving love? Enduring grief activates brain's reward center. Neuroimage 42: 969-972. doi: 10.1016/j.neuroimage.2008.04.256
|
| [216] |
Shear K, Frank E, Houck PR, et al. (2005) Treatment of complicated grief: A randomized controlled trial. J Am Med Assoc 293: 2601-2608. doi: 10.1001/jama.293.21.2601
|
Figures(1) / Tables(1)

Robert A. Moss. Psychotherapy in pain management: New viewpoints and treatment targets based on a brain theory[J]. AIMS Neuroscience, 2020, 7(3): 194-207. doi: 10.3934/Neuroscience.2020013







 DownLoad:
DownLoad: