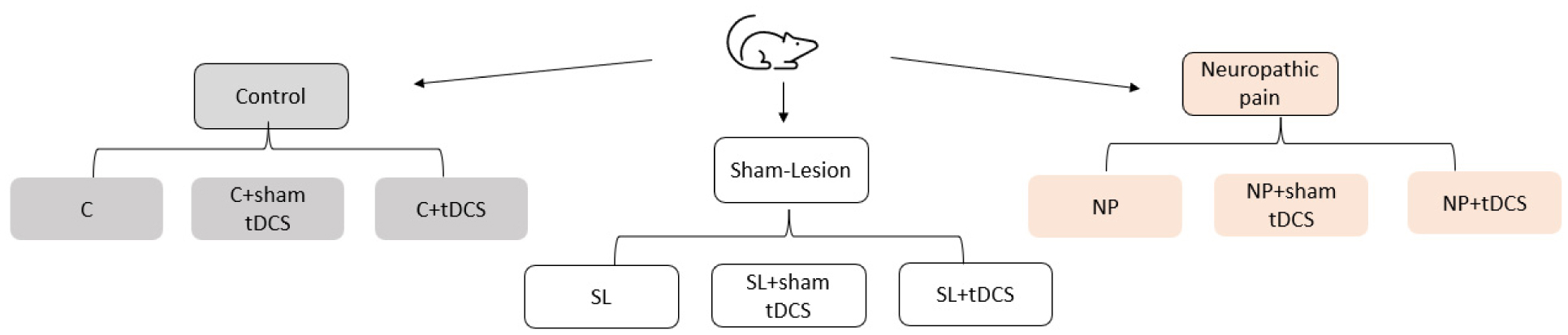
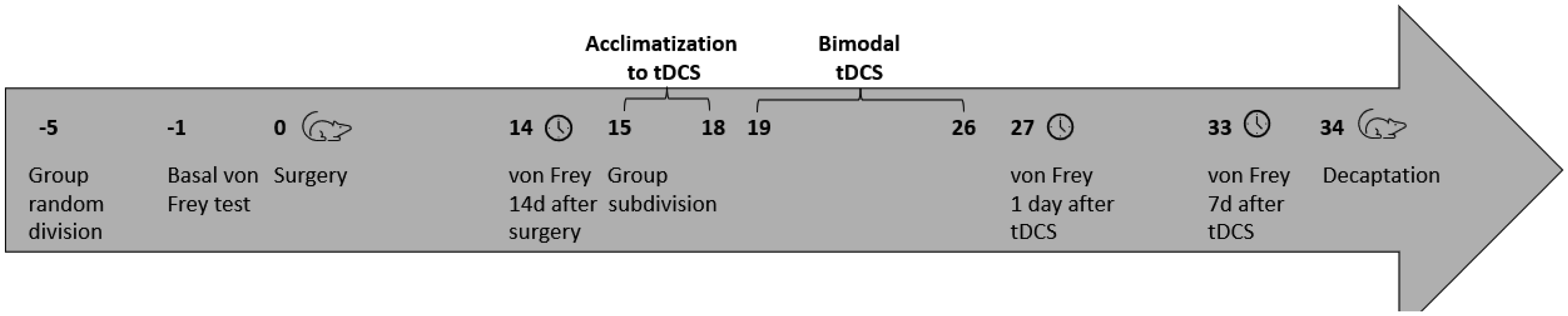
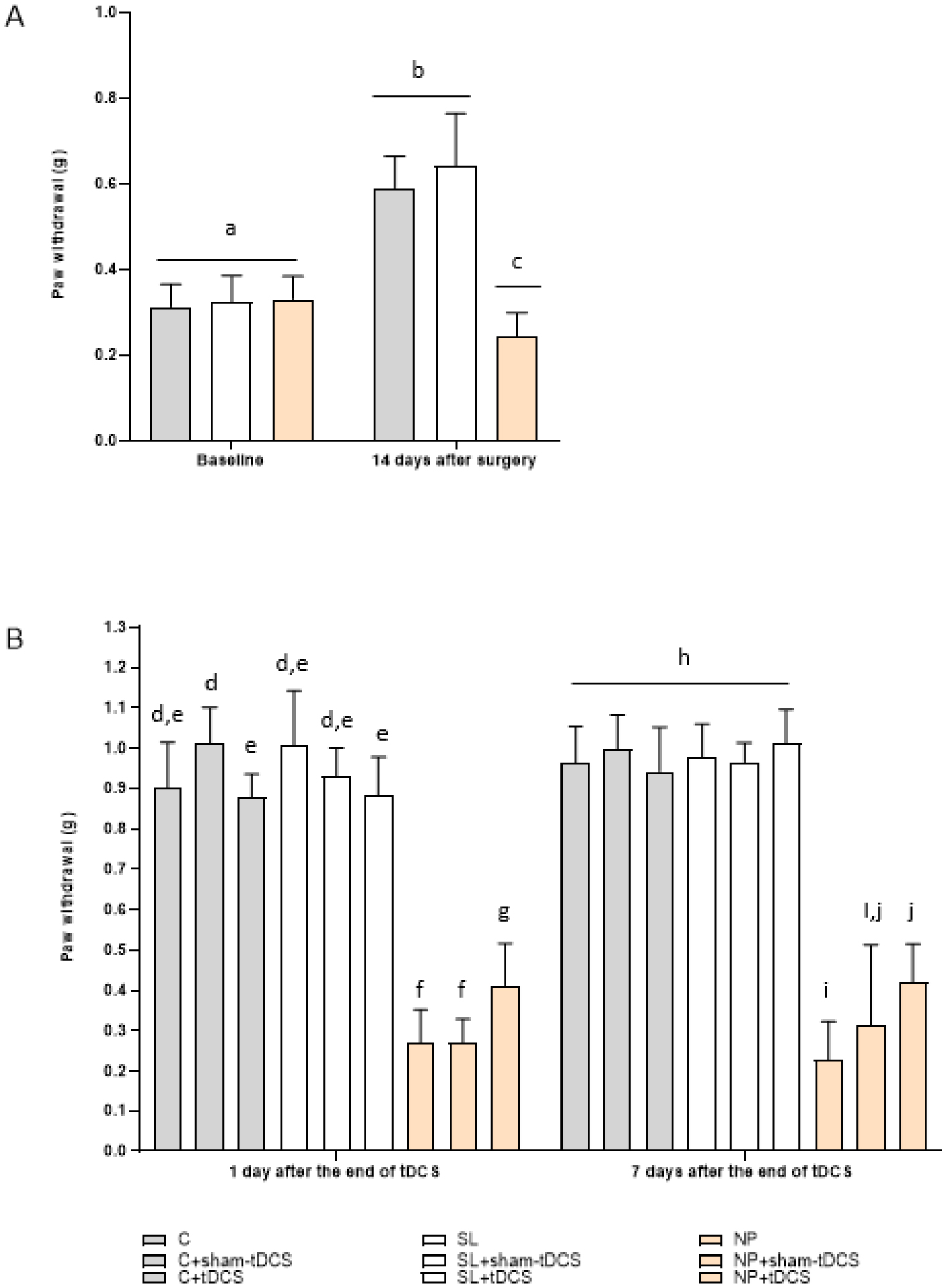
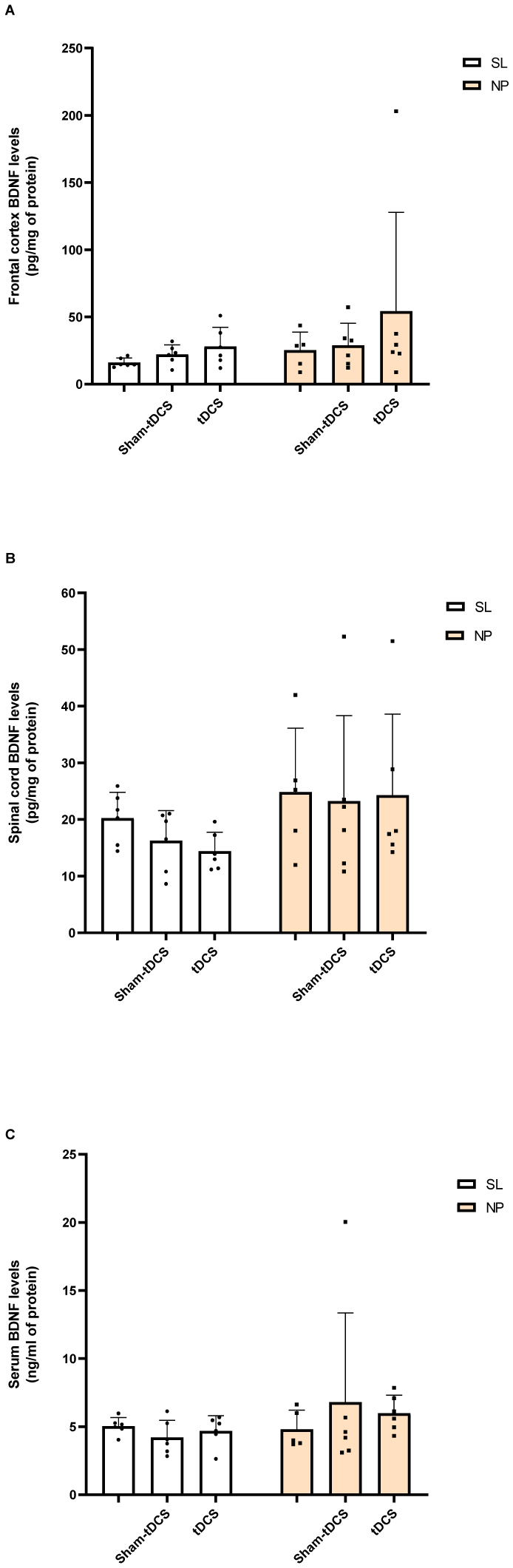
Neuropathic pain (NP) is caused by a lesion that triggers pain chronification and central sensitization and it can develop in a different manner, dependent of age. Recent studies have demonstrated the efficacy of transcranial direct current stimulation (tDCS) for treating NP. Then, we aimed to investigate the effects of tDCS and BDNF levels in neuropathic pain rats in development, with 30 days old in the beginning of experiments. Eight-five male Wistar rats were subjected to chronic constriction injury. After establishment of NP, bimodal tDCS was applied to the rats for eight consecutive days, for 20 minutes each session. Subsequently, nociceptive behavior was assessed at baseline, 14 days after surgery, 1 day and 7 days after the end of tDCS. The rats were sacrificed 8 days after the last session of tDCS. An increase in the nociceptive threshold was observed in rats in development 1 day after the end of tDCS (short-term effect), but this effect was not maintained 7 days after the end of tDCS (long-term effect). Furthermore, brain derived neurotrophic factor (BDNF) levels were analyzed in the frontal cortex, spinal cord and serum using ELISA assays. The neuropathic pain model showed an effect of BDNF in the spinal cord of rats in development. There were no effects of BNDF levels of pain or tDCS in the frontal cortex or serum. In conclusion, tDCS is an effective technique to relieve nociceptive behavior at a short-term effect in neuropathic pain rats in development, and BDNF levels were not altered at long-term effect.
Citation: Priscila Centeno Crespo, Leo Anderson Meira Martins, Otávio Garcia Martins, Clara Camacho Dos Reis, Ricardo Netto Goulart, Andressa de Souza, Liciane Fernandes Medeiros, Vanessa Leal Scarabelot, Giovana Duzzo Gamaro, Sabrina Pereira Silva, Marcos Roberto de Oliveira, Iraci Lucena da Silva Torres, Izabel Cristina Custódio de Souza. Short-term effectiveness of transcranial direct current stimulation in the nociceptive behavior of neuropathic pain rats in development[J]. AIMS Neuroscience, 2023, 10(4): 433-446. doi: 10.3934/Neuroscience.2023032
Neuropathic pain (NP) is caused by a lesion that triggers pain chronification and central sensitization and it can develop in a different manner, dependent of age. Recent studies have demonstrated the efficacy of transcranial direct current stimulation (tDCS) for treating NP. Then, we aimed to investigate the effects of tDCS and BDNF levels in neuropathic pain rats in development, with 30 days old in the beginning of experiments. Eight-five male Wistar rats were subjected to chronic constriction injury. After establishment of NP, bimodal tDCS was applied to the rats for eight consecutive days, for 20 minutes each session. Subsequently, nociceptive behavior was assessed at baseline, 14 days after surgery, 1 day and 7 days after the end of tDCS. The rats were sacrificed 8 days after the last session of tDCS. An increase in the nociceptive threshold was observed in rats in development 1 day after the end of tDCS (short-term effect), but this effect was not maintained 7 days after the end of tDCS (long-term effect). Furthermore, brain derived neurotrophic factor (BDNF) levels were analyzed in the frontal cortex, spinal cord and serum using ELISA assays. The neuropathic pain model showed an effect of BDNF in the spinal cord of rats in development. There were no effects of BNDF levels of pain or tDCS in the frontal cortex or serum. In conclusion, tDCS is an effective technique to relieve nociceptive behavior at a short-term effect in neuropathic pain rats in development, and BDNF levels were not altered at long-term effect.

| [1] | International association for the study of pain. What is neuropathic pain?. Global year against neuropathic pain, USA (2014) . Avalaible from: https://www.iasp-pain.org |
| [2] |
Cioato SG, Medeiros LF, Marques Filho PR, et al. (2016) Long-Lasting Effect of Transcranial Direct Current Stimulation in the Reversal of Hyperalgesia and Cytokine Alterations Induced by the Neuropathic Pain Model. Brain Stimul 2: 209-217. https://doi.org/10.1016/j.brs.2015.12.001 
|
| [3] |
Vega-Avelaira D, Géranton SM, Fitzgerald M (2009) Differential regulation of immune responses and macrophage/neuron interactions in the dorsal root ganglion in young and adult rats following nerve injury. Mol Pain 5: 1-17. https://doi.org/10.1186/1744-8069-5-70
|
| [4] | Austin PJ, Wu A, Moalem-Taylor G (2012) Chronic constriction of the sciatic nerve and pain hypersensitivity testing in rats. J Vis Exp : e3393. https://doi.org/10.3791/3393 |
| [5] | Boldt I, Eriks-Hoogland I, Brinkhof MW, et al. (2014) Non-pharmacological interventions for chronic pain in people with spinal cord injury. Cochrane Database Syst Rev 11: CD009177. https://doi.org/10.1002/14651858.CD009177.pub2 |
| [6] |
Antal A, Terney D, Kühnl S, et al. (2010) Anodal Transcranial Direct Current Stimulation of the Motor Cortex Ameliorates Chronic Pain and Reduces Short Intracortical Inhibition. J Pain Symptom Manage 39: 890-903. https://doi.org/10.1016/j.jpainsymman.2009.09.023
|
| [7] |
Liu A, Bryant A, Jefferson A, et al. (2016) Exploring the efficacy of a 5-day course of transcranial direct current stimulation (TDCS) on depression and memory function in patients with well-controlled temporal lobe epilepsy. Epilepsy Behav 55: 11-20. https://doi.org/10.1016/j.yebeh.2015.10.032
|
| [8] |
Dimov LF, Franciosi AC, Pinheiro Campos AC, et al. (2016) Top-Down Effect of Direct Current Stimulation on the Nociceptive Response of Rats. PLoS One 11: e0153506. https://doi.org/10.1371/journal.pone.0153506
|
| [9] |
Roche N, Lackmy A, Achache V, et al. (2011) Effects of anodal transcranial direct current stimulation over the leg motor area on lumbar spinal network excitability in healthy subjects. J Physiol 589: 2813-2826. https://doi.org/10.1113/jphysiol.2011.205161
|
| [10] | Schestatsky P, Morales-Quezada L, Fregni F (2013) Simultaneous EEG monitoring during transcranial direct current stimulation. J Vis Exp 76: e50426. https://doi.org/10.3791/50426 |
| [11] | Palm U, Chalah MA, Padberg F, et al. (2016) Effects of transcranial random noise stimulation (tRNS) on affect, pain and attention in multiple sclerosis. Restor Neurol Neurosci 34: 189-199. https://doi.org/10.3233/RNN-150557 |
| [12] |
Filho PRM, Vercelino R, Cioato SG, et al. (2016) Transcranial direct current stimulation (tDCS) reverts behavioral alterations and brainstem BDNF level increase induced by neuropathic pain model: Long-lasting effect. Prog Neuro-Psychopharmacology Biol Psychiatry 64: 44-51. https://doi.org/10.1016/j.pnpbp.2015.06.016
|
| [13] | DaSilva AF, Volz MS, Bikson M, et al. (2011) Electrode positioning and montage in transcranial direct current stimulation. J Vis Exp 51: e2744. https://doi.org/10.3791/2744 |
| [14] |
Spezia Adachi LN, Caumo W, Laste G, et al. (2012) Reversal of chronic stress-induced pain by transcranial direct current stimulation (tDCS) in an animal model. Brain Res 1489: 17-26. https://doi.org/10.1016/j.brainres.2012.10.009
|
| [15] |
Fujiyama H, Hyde J, Hinder MR, et al. (2014) Delayed plastic responses to anodal tDCS in older adults. Front Aging Neurosci 6: 115. https://doi.org/10.3389/fnagi.2014.00115
|
| [16] |
Ayache SS, Palm U, Chalah MA, et al. (2016) Prefrontal tDCS decreases pain in patients with multiple sclerosis. Front Neurosci 10: 147. https://doi.org/10.3389/fnins.2016.00147
|
| [17] |
Knotkova H, Portenoy RK, Cruciani RA (2013) Transcranial direct current stimulation (tDCS) relieved itching in a patient with chronic neuropathic pain. Clin J Pain 29: 621-622. https://doi.org/10.1097/AJP.0b013e31826b1329
|
| [18] |
Volz MS, Farmer A, Siegmund B (2016) Reduction of chronic abdominal pain in patients with inflammatory bowel disease through transcranial direct current stimulation: a randomized controlled. Pain 157: 429-437. https://doi.org/10.1097/j.pain.0000000000000386
|
| [19] |
Khan N, Smith MT (2015) Neurotrophins and neuropathic pain: Role in pathobiology. Molecules 20: 10657-10688. https://doi.org/10.3390/molecules200610657
|
| [20] |
Oh H, Lewis DA, Sibille E (2016) The Role of BDNF in Age-Dependent Changes of Excitatory and Inhibitory Synaptic Markers in the Human Prefrontal Cortex. Neuropsychopharmacology 41: 3080-3091. https://doi.org/10.1038/npp.2016.126
|
| [21] |
Lopes BC, Medeiros LF, Silva de Souza V, et al. (2020) Transcranial direct current stimulation combined with exercise modulates the inflammatory profile and hyperalgesic response in rats subjected to a neuropathic pain model: Long-term effects. Brain Stimul 13: 774-782. https://doi.org/10.1016/j.brs.2020.02.025
|
| [22] |
Lopes BC, Medeiros LF, Stein DJ, et al. (2021) tDCS and exercise improve anxiety-like behavior and locomotion in chronic pain rats via modulation of neurotrophins and inflammatory mediators. Behav Brain Res 404: 113173. https://doi.org/10.1016/j.bbr.2021.113173
|
| [23] |
Santos DS, Medeiros LF, Stein DJ, et al. (2021) Bimodal transcranial direct current stimulation reduces alcohol consumption and induces long-term neurochemical changes in rats with neuropathic pain. Neurosci Lett 759: 136014. https://doi.org/10.1016/j.neulet.2021.136014
|
| [24] |
Silva SP, Martins OG, Medeiros LF, et al. (2022) Evidence of Anti-Inflammatory Effect of Transcranial Direct Current Stimulation in a CFA-Induced Chronic Inflammatory Pain Model in Wistar Rats. Neuroimmunomodulation 29: 500-514. https://doi.org/10.1159/000520581
|
| [25] |
Netto CA, Siegfried B, Izquierdo I (1987) Analgesia induced by exposure to a novel environment in rats: Effect of concurrent and post-training stressful stimulation. Behav Neural Biol 48: 304-309. https://doi.org/10.1016/S0163-1047(87)90850-8
|
| [26] |
Vivancos GG, Verri WA, Cunha TM, et al. (2004) An electronic pressure-meter nociception paw test for rats. Brazilian J Med Biol Res 37: 391-399. https://doi.org/10.1590/S0100-879X2004000300017
|
| [27] |
Bennett GJ, Xie YK (1988) A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 33: 87-107. https://doi.org/10.1016/0304-3959(88)90209-6
|
| [28] |
Santos DS, Lopes BC, Medeiros LF, et al. (2020) Transcranial Direct Current Stimulation (tDCS) Induces Analgesia in Rats with Neuropathic Pain and Alcohol Abstinence. Neurochem Res 45: 2653-2663. https://doi.org/10.1007/s11064-020-03116-w
|
| [29] |
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254. https://doi.org/10.1016/0003-2697(76)90527-3
|
| [30] |
Nunes EA, Medeiros LF, de Freitas JS, et al. (2016) Morphine Exposure during Early Life Alters Thermal and Mechanical Thresholds in Rats. Int J Dev Neurosci 60: 78-85. https://doi.org/10.1016/j.ijdevneu.2016.12.008
|
| [31] |
Boada MD, Gutierrez S, Aschenbrenner CA, et al. (2015) Nerve injury induces a new profile of tactile and mechanical nociceptor input from undamaged peripheral afferents. J Neurophysiol 113: 100-109. https://doi.org/10.1152/jn.00506.2014
|
| [32] |
Lu VB, Biggs JE, Stebbing MJ, et al. (2009) Brain-derived neurotrophic factor drives the changes in excitatory synaptic transmission in the rat superficial dorsal horn that follow sciatic nerve injury. J Physiol 587: 1013-1032. https://doi.org/10.1113/jphysiol.2008.166306
|
| [33] |
Howard RF, Walker SM, Mota MP, et al. (2005) The ontogeny of neuropathic pain: postnatal onset of mechanical allodynia in rat spared nerve injury (SNI) and chronic constriction injury (CCI) models. Pain 115: 382-389. https://doi.org/10.1016/j.pain.2005.03.016
|
| [34] |
Centeno Crespo P, Anderson Meira Martins L, Camacho Dos Reis C, et al. (2023) Transcranial direct current stimulation effects in the pain threshold and in oxidative stress parameters of neuropathic pain rats. Neurosci Lett 11: 137179. https://doi.org/10.1016/j.neulet.2023.137179
|
| [35] |
Csifcsak G, Antal A, Hillers F, et al. (2009) Modulatory effects of transcranial direct current stimulation on laser-evoked potentials. Pain Med 10: 122-132. https://doi.org/10.1111/j.1526-4637.2008.00508.x
|
| [36] |
Vaseghi B, Zoghi M, Jaberzadeh S (2015) The effects of anodal-tDCS on corticospinal excitability enhancement and its after-effects: conventional vs. unihemispheric concurrent dual-site stimulation. Front Hum Neurosci 9: 1-13. https://doi.org/10.3389/fnhum.2015.00533
|
| [37] |
Coull JA, Beggs S, Boudreau D, et al. (2005) BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nature 438: 1017-1021. https://doi.org/10.1038/nature04223
|
| [38] |
Soltész F, Suckling J, Lawrence P, et al. (2014) Identification of BDNF sensitive electrophysiological markers of synaptic activity and their structural correlates in healthy subjects using a genetic approach utilizing the functional BDNF Val66Met polymorphism. PLoS One 9: e95558. https://doi.org/10.1371/journal.pone.0095558
|
| [39] |
Cheeran B, Talelli P, Mori F, et al. (2008) A common polymorphism in the brain derived neurotrophic factor gene (BDNF) modulates human cortical plasticity and the response to rTMS. J Physiol 586: 5717-5725. https://doi.org/10.1113/jphysiol.2008.159905
|
| [40] |
Kuo MF, Paulus W, Nitsche MA (2014) Therapeutic effects of non-invasive brain stimulation with direct currents (tDCS) in neuropsychiatric diseases. Neuroimage 85: 948-960. https://doi.org/10.1016/j.neuroimage.2013.05.117
|
| [41] |
Pirulli C, Fertonani A, Miniussi C (2013) The role of timing in the induction of neuromodulation in perceptual learning by transcranial electric stimulation. Brain Stimul 6: 683-689. https://doi.org/10.1016/j.brs.2012.12.005
|
| [42] |
Boudes M, Menigoz A (2009) Non-neuronal BDNF, a key player in development of central sensitization and neuropathic pain. J Physiol 587: 2111-2112. https://doi.org/10.1113/jphysiol.2009.172130
|
Figures(4)

Priscila Centeno Crespo, Leo Anderson Meira Martins, Otávio Garcia Martins, Clara Camacho Dos Reis, Ricardo Netto Goulart, Andressa de Souza, Liciane Fernandes Medeiros, Vanessa Leal Scarabelot, Giovana Duzzo Gamaro, Sabrina Pereira Silva, Marcos Roberto de Oliveira, Iraci Lucena da Silva Torres, Izabel Cristina Custódio de Souza. Short-term effectiveness of transcranial direct current stimulation in the nociceptive behavior of neuropathic pain rats in development[J]. AIMS Neuroscience, 2023, 10(4): 433-446. doi: 10.3934/Neuroscience.2023032







 DownLoad:
DownLoad: