Citation: Kelly F. Austin, Megan O. Bellinger, Priyokti Rana. Anthropogenic forest loss and malaria prevalence: a comparative examination of the causes and disease consequences of deforestation in developing nations[J]. AIMS Environmental Science, 2017, 4(2): 217-231. doi: 10.3934/environsci.2017.2.217

| [1] | World Health Organization (WHO), World Malaria Report 2016, 2016. Available from: http://apps.who.int/iris/bitstream/10665/252038/1/9789241511711-eng.pdf?ua=1. |
| [2] |
Norris D (2004) Mosquito-borne Diseases as a Consequence of Land Use Change. EcoHealth 1: 19-24. doi: 10.1007/s10393-004-0008-7

|
| [3] |
Lewis S, Maslin M (2015) Defining the Anthropocene. Nature 519: 171-180. doi: 10.1038/nature14258
|
| [4] | Food and Agriculture Organization (FAO) of the United Nations, Global Forest Resources Assessment 2015, 2015. Available from: http://www.fao.org/3/a-i4808e.pdf. |
| [5] |
Ernst KC, Lindblade KA, Koech D, et al. (2009) Environmental, socio-demographic and behavioural determinants of malaria risk in the western Kenyan highlands: a case-control study. Trop Med Int Health 14: 1258-1265. doi: 10.1111/j.1365-3156.2009.02370.x
|
| [6] | Vittor AY, Pan W, Gilman RH, et al. (2009) Linking deforestation to malaria in the Amazon: characterization of the breeding habitat of the principal malaria vector, Anopheles darlingi. Am J Trop Med Hyg 81: 5-12. |
| [7] |
Hanandita W, Tampubolon G (2016) Geography and social distribution of malaria in Indonesian Papua: a cross-sectional study. Int J Health Geogr 15: 13. doi: 10.1186/s12942-016-0043-y
|
| [8] |
Wang X, Zhou G, Zhong D, et al. (2016) Life-table studies revealed significant effects of deforestation on the development and survivorship of Anopheles minimus larvae. Parasite vector 9: 323. doi: 10.1186/s13071-016-1611-5
|
| [9] | Vittor AY, Gilman RH, Tielsch J, et al. (2006) The effect of deforestation on the human-biting rate of Anopheles darlingi, the primary vector of falciparum malaria in the Peruvian Amazon. Am J Trop Med Hyg 74: 3-11. |
| [10] |
Manh BH, Clements ACA, Thieu NQ, et al. (2011) Social and environmental determinants of malaria in space and time in Viet Nam. Int J Parasitol 41: 109-116. doi: 10.1016/j.ijpara.2010.08.005
|
| [11] | World Health Organization (WHO), World Malaria Report 2015, 2015. Available from: http://www.who.int/malaria/publications/world-malaria-report-2015/report/en/. |
| [12] | Packard RM, The making of a tropical disease: a short history of malaria. JHU Press, 2007. |
| [13] |
Stratton L, O'Neill MS, Kruk ME, et al. (2008) The persistent problem of malaria: Addressing the fundamental causes of a global killer. Soc Sci Med 67: 854-862. doi: 10.1016/j.socscimed.2008.05.013
|
| [14] |
Austin KF, Noble MD, Mejia MT (2014) Gendered vulnerabilities to a neglected disease: A comparative investigation of the effect of women's legal economic rights and social status on malaria rates. Int J Comp Sociol 55: 204-228. doi: 10.1177/0020715214543158
|
| [15] |
Brady D, Kaya Y, Beckfield J (2007) Reassessing the effect of economic growth on well-being in less-developed countries, 1980–2003. Stud Comp Int Develop 42: 1-35. doi: 10.1007/s12116-007-9003-7
|
| [16] |
Burroway R (2010) Schools against AIDS: Secondary school enrollment and cross-national disparities in AIDS death rates. Soc Probl 57: 398-420. doi: 10.1525/sp.2010.57.3.398
|
| [17] |
Shen C, Williamson JB (2001) Accounting for cross-national differences in infant mortality decline (1965–1991) among less developed countries: Effects of women's status, economic dependency, and state strength. Soc Indic Res 53: 257-288. doi: 10.1023/A:1007190612314
|
| [18] |
Williamson JB, Boehmer U (1997) Female life expectancy, gender stratification, health status, and level of economic development: A cross-national study of less developed countries. Soc Sci Med 45: 305-317. doi: 10.1016/S0277-9536(96)00346-2
|
| [19] | Mayxay M, Pukrittayakamee S, Newton PN, et al. (2004) Mixed-species malaria infections in humans. Trends Parasitol 20: 233-240. |
| [20] |
Neafsey DE, Waterhouse RM, Abai MR, et al. (2015) Highly evolvable malaria vectors: the genomes of 16 Anopheles mosquitoes. Science 347: 1258522. doi: 10.1126/science.1258522
|
| [21] | Bates I, Fenton C, Gruber J, et al. (2004) Vulnerability to malaria, tuberculosis, and HIV/AIDS infection and disease. Part II: determinants operating at environmental and institutional level. Lancet Infect Diseases 4: 368-375. |
| [22] | Hastings IM, Ward SA (2005) Coartem (artemether-lumefantrine) in Africa: the beginning of the end? J Infect Diseases 192: 1303-1304. |
| [23] | Shah S, The Fever, Picador, 2010. |
| [24] |
Sachs J, Malaney P (2002) The economic and social burden of malaria. Nature 415: 680-685. doi: 10.1038/415680a
|
| [25] |
Lowassa A, Mazigo HD, Mahande AM, et al. (2012) Social economic factors and malaria transmission in Lower Moshi, northern Tanzania. Parasite vector 5: 129. doi: 10.1186/1756-3305-5-129
|
| [26] |
Williams PCM, Martina A, Cumming RG, et al. (2009) Malaria prevention in Sub-Saharan Africa: a field study in rural Uganda. J Commun Health 34: 288-294. doi: 10.1007/s10900-009-9151-y
|
| [27] |
Dike N, Onwujekwe O, Ojukwu J, et al. (2006) Influence of education and knowledge on perceptions and practices to control malaria in Southeast Nigeria. Soc Sci Med 63: 103-106. doi: 10.1016/j.socscimed.2005.11.061
|
| [28] |
Williams HA, Jones COH (2004) A critical review of behavioral issues related to malaria control in sub-Saharan Africa: what contributions have social scientists made? Soc Sci Med 59: 501-523. doi: 10.1016/j.socscimed.2003.11.010
|
| [29] | Blanford JI, Blanford S, Crane RG, et al. (2013) Implications of temperature variation for malaria parasite development across Africa. Sci Rep 3: 1300. |
| [30] | Pattanayak S, Dickinson K, Corey C, et al. (2006) Deforestation, malaria, and poverty: a call for transdisciplinary research to support the design of cross-sectoral policies. Sustainability: Science, Practice, Policy 2(2). |
| [31] |
Pimentel D, Cooperstein S, Randell H, et al. (2007) Ecology of increasing diseases: population growth and environmental degradation. Hum Ecol 35: 653-668. doi: 10.1007/s10745-007-9128-3
|
| [32] |
Patz JA, Graczyk TK, Geller N, et al. (2000) Effects of environmental change on emerging parasitic diseases. Int J Parasitol 30: 1395-1405. doi: 10.1016/S0020-7519(00)00141-7
|
| [33] |
Myers SS, Gaffikin L, Golden CD, et al. (2013) Human health impacts of ecosystem alteration. Proc Nat Acad Sci USA 110: 18753-18760. doi: 10.1073/pnas.1218656110
|
| [34] |
Laporta GZ, de Prado PIKL, Kraenkel RA, et al. (2013) Biodiversity can help prevent malaria outbreaks in tropical forests. PLoS Negl Trop Dis 7: e2139. doi: 10.1371/journal.pntd.0002139
|
| [35] | Yasuoka J, Levins R (2007) Impact of deforestation and agricultural development on anopheline ecology and malaria epidemiology. Am J Trop Med Hyg 76: 450-460. |
| [36] | World Bank, World development report: Agriculture for development, 2008. |
| [37] |
Bensel T (2008) Fuelwood, deforestation, and land degradation: 10 years of evidence from Cebu Province, the Philippines. Land Degrad Dev 19: 587-605. doi: 10.1002/ldr.862
|
| [38] |
Jorgenson AK, Burns TJ (2007) Effects of Rural and Urban Population Dynamics and National Development on Deforestation in Less-Developed Countries, 1990–2000. Sociol Inq 77: 460-482. doi: 10.1111/j.1475-682X.2007.00200.x
|
| [39] | Bollen K, Structural Equations with Latent Variables, New York: John Wiley and Sons, Inc, 1989. |
| [40] | Kaplan D, Structural equation modeling: Foundations and extensions. Sage Publications, 2008. |
| [41] | Muthén LK, Muthén BO, Mplus User's Guide, 5th ed., Los Angeles: Muthén and Muthén, 2007. |
| [42] | Arbuckle JL (1996) Full information estimation in the presence of incomplete data. Advanced structural equation modeling: Issues and technique 243: 277. |
| [43] | World Bank, World Databank, World Development Indicators, 2016. Available from: http://databank.worldbank.org/data/views/variableSelection/selectvariables.aspx?source=world-development-indicators#. |
| [44] |
Austin KF, McKinney LA (2012) Disease, war, hunger, and deprivation: A cross-national investigation of the determinants of life expectancy in less-developed and sub-Saharan African nations. Sociol Perspect 55: 421-447. doi: 10.1525/sop.2012.55.3.421
|
| [45] | Shandra CL, Shandra JM, London B (2011) World bank structural adjustment, water, and sanitation: A cross-national analysis of child mortality in Sub-Saharan Africa. Organ Environ 24: 107-129. |
| [46] |
Austin KF, McKinney LA, Thompson G (2012) Agricultural Trade Dependency and the Threat of Starvation: A Cross-National Analysis of Hunger as Unequal Exchange. Int J Sociol 42: 68-89. doi: 10.2753/IJS0020-7659420204
|
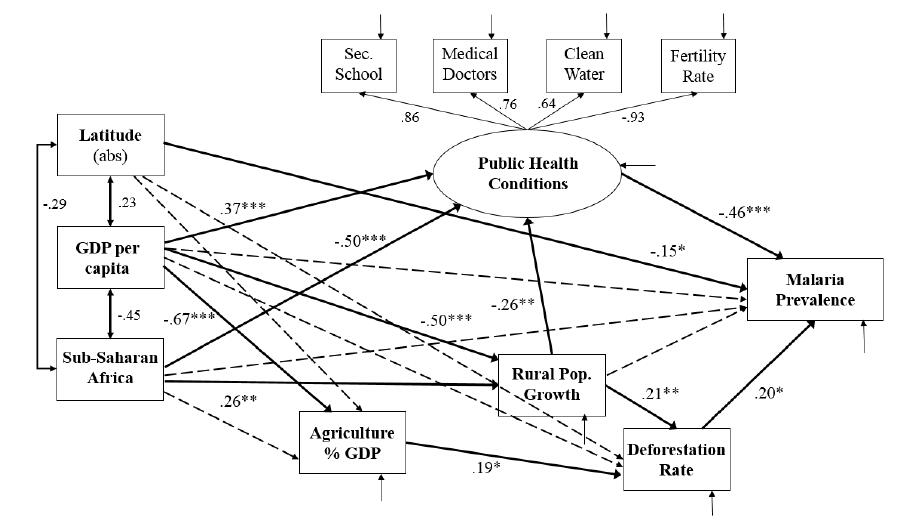
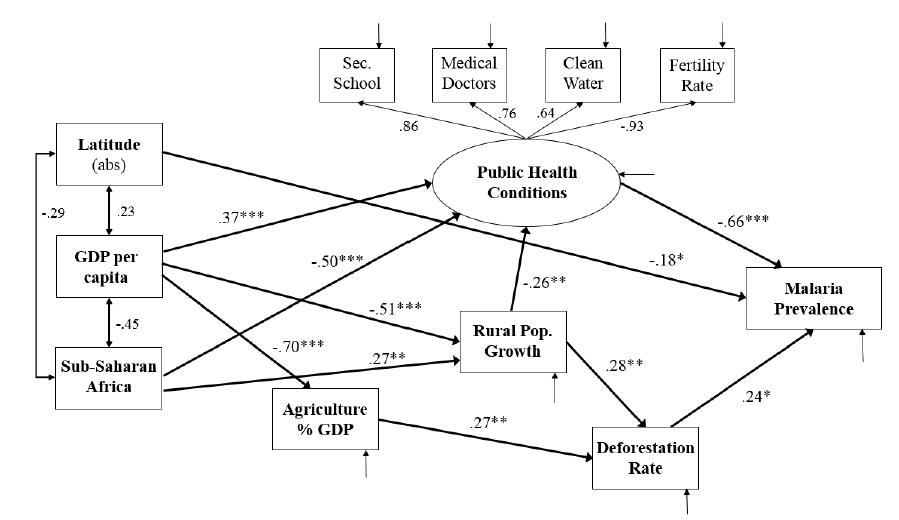
Figures(2) / Tables(3)

Kelly F. Austin, Megan O. Bellinger, Priyokti Rana. Anthropogenic forest loss and malaria prevalence: a comparative examination of the causes and disease consequences of deforestation in developing nations[J]. AIMS Environmental Science, 2017, 4(2): 217-231. doi: 10.3934/environsci.2017.2.217







 DownLoad:
DownLoad: