Citation: Robert Russell Monteith Paterson. Depletion of Indonesian oil palm plantations implied from modeling oil palm mortality and Ganoderma boninense rot under future climate[J]. AIMS Environmental Science, 2020, 7(5): 366-379. doi: 10.3934/environsci.2020024

| [1] | Paterson RRM, Lima N (2018) Climate change affecting oil palm agronomy, and oil palm cultivation increasing climate change, require amelioration. Ecol Evol 8: 452-461. |
| [2] | Paterson RRM (2019) Ganoderma boninense disease of oil palm is expected to significantly reduce production after 2050 in Sumatra if projected climate change occurs. Microorganisms 7: 24. |
| [3] | Paterson RRM (2019) Ganoderma boninense disease deduced from simulation modelling with large data sets of future Malaysian oil palm climate. Phytoparasitica 47: 255-262. |
| [4] | Paterson RRM (2020). Oil palm survival under climate change in Kalimantan and alternative SE Asian palm oil countries with future basal stem rot assessments. Forest Pathol 50: e12604. |
| [5] | Paterson RRM (2020) Future scenarios for oil palm mortality and infection by Phytophthora palmivora in Colombia, Ecuador and Brazil, extrapolated to Malaysia and Indonesia. Phytoparasitica 48: 513-523. |
| [6] | Sarkar SK, Begum RA, Pereira JJ (2020) Impacts of climate change on oil palm production in Malaysia. Environ Sci Pollut R 27: 9760-9770. |
| [7] | Corley RHV, Tinker PB (2015) The Oil Palm, Wiley Blackwell. |
| [8] | Lam YW, Kulak M, Sim S, et al. (2019) Greenhouse gas footprints of palm oil production in Indonesia over space and time. Sci Total Environ 688: 827-837. |
| [9] | Rianto B (2010) Overview of palm oil industry in Indonesia. Pricewaterhouse Coopers Indonesia. Available from: https://www.pwc.com/id/en/publications/assets/palm-oil-plantation.pdf |
| [10] | Paterson RRM, Kumar L, Taylor S, et al. (2015) Future climate effects on suitability for growth of oil palms in Malaysia and Indonesia. Sci Report 5. |
| [11] | Paterson RRM, Kumar L, Shabani F, et al. (2017) World climate suitability projections to 2050 and 2100 for growing oil palm. J Agric Sci 155: 689-702. |
| [12] | Dislich C, Keyel AC, Salecker J, et al. (2017) Wiegand, K. A review of the ecosystem functions in oil palm plantations, using forests as a reference system. Biol Rev 49: 1539-1569. |
| [13] | Meijaard E, Garcia-Ulloa J, Sheil D, et al. (2018) Oil palm and biodiversity: A situation analysis by the IUCN Oil Palm Task Force. 2018. |
| [14] | Kadandale S, Marten R, Smith R (2019) The palm oil industry and noncommunicable diseases. Bull World Healt Organ 97: 118-128. |
| [15] | Gibb R, Redding DW, Chin KQ, et al. (2020). Zoonotic host diversity increases in human-dominated ecosystems. Nature 584: 398-402. |
| [16] | Paterson RRM, Sariah M, Lima N (2013) How will climate change affect oil palm fungal diseases? Crop Prot 46: 113-120. |
| [17] | Zhou LW, Cao Y, Wu SH, et al. (2015) Global diversity of the Ganoderma lucidum complex (Ganodermataceae, Polyporales) inferred from morphology and multilocus phylogeny. Phytochem 114: 7-15. |
| [18] | Merciere M, Boulord R, Carasco-Lacombe C, et al. (2017) About Ganoderma boninense in oil palm plantations of Sumatra and peninsular Malaysia: Ancient population expansion, extensive gene flow and large scale dispersion ability. Fungal Biol 121: 529-540. |
| [19] | Paterson R R M, Lima N. Ecology and biotechnology of thermophilic fungi on crops under global warming[M]//Fungi in Extreme Environments: Ecological Role and Biotechnological Significance. Springer, Cham. 2019: 81-96. |
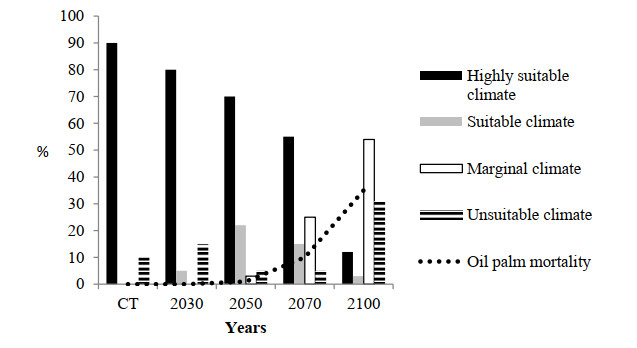
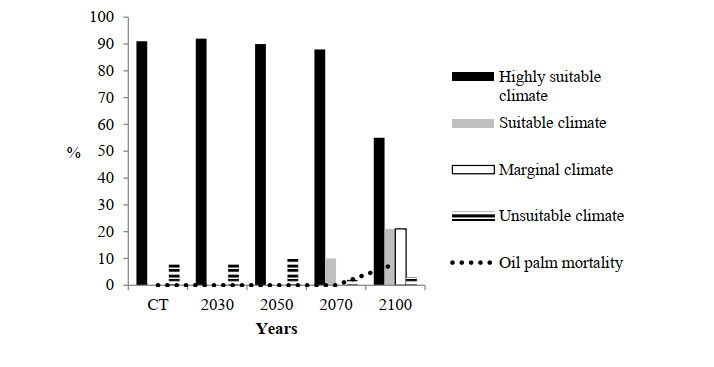
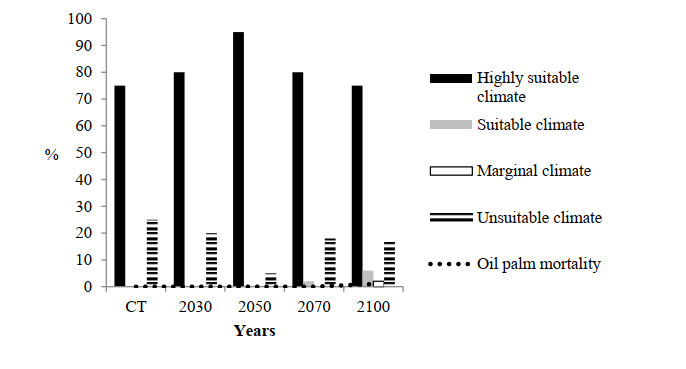
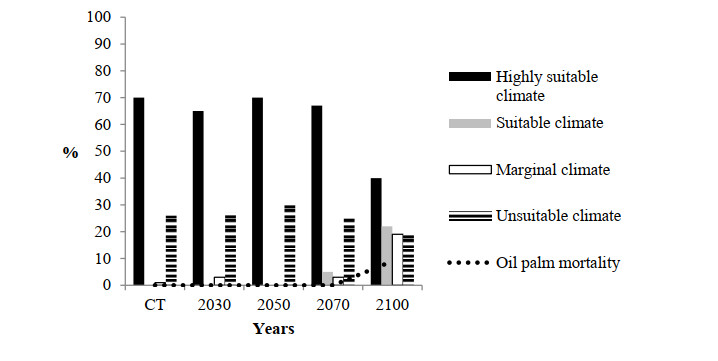
Figures(10)

Robert Russell Monteith Paterson. Depletion of Indonesian oil palm plantations implied from modeling oil palm mortality and Ganoderma boninense rot under future climate[J]. AIMS Environmental Science, 2020, 7(5): 366-379. doi: 10.3934/environsci.2020024







 DownLoad:
DownLoad: