Citation: Frank Tietz. Phase relations of NASICON materials and compilation of the quaternary phase diagram Na2O-P2O5-SiO2-ZrO2[J]. AIMS Materials Science, 2017, 4(6): 1305-1318. doi: 10.3934/matersci.2017.6.1305

| [1] | Larcher D, Tarascon JM (2015) Towards greener and more sustainable batteries for electrical energy storage. Nat Chem 7: 19–29. |
| [2] | Doughty DH, Butler PC, Akhil AA, et al. (2010) Batteries for Large-Scale Stationary Electrical Energy Storage. Electrochem Soc Interface 19: 49–53. |
| [3] |
Benato R, Cosciani N, Crugnola G, et al. (2015) Sodium nickel chloride battery technology for large-scale stationary storage in the high voltage network. J Power Sources 293: 127–136. doi: 10.1016/j.jpowsour.2015.05.037

|
| [4] |
Lu X, Xia G, Lemmon JP, et al. (2010) Advanced materials for sodium-beta alumina batteries: Status, challenges and perspectives. J Power Sources 195: 2431–2442. doi: 10.1016/j.jpowsour.2009.11.120
|
| [5] |
Hong HYP (1976) Crystal Structure and Crystal Chemistry in the System Na1+xZr2SixP3−xO12. Mater Res Bull 11: 173–182. doi: 10.1016/0025-5408(76)90073-8
|
| [6] |
Goodenough JB, Hong HYP, Kafalas JA (1976) Fast Na+-Ion Transport in Skeleton Structures. Mater Res Bull 11: 203–220. doi: 10.1016/0025-5408(76)90077-5
|
| [7] |
Kim J, Jo SH, Bhavaraju S, et al. (2015) Low temperature performance of sodium–nickel chloride batteries with NaSICON solid electrolyte. J Electroanal Chem 759: 201–206. doi: 10.1016/j.jelechem.2015.11.022
|
| [8] |
May GJ, Hooper A (1978) The effect of microstructure and phase composition on the ionic conductivity of magnesium-doped sodium-beta-alumina. J Mater Sci 13: 1480–1486. doi: 10.1007/BF00553202
|
| [9] |
Clearfield A, Guerra R, Oskarsson A, et al. (1983) Preparation of Sodium Zirconium Phosphates of the Type Na1+4xZr2−x(PO4)3. Mater Res Bull 18: 1561–1567. doi: 10.1016/0025-5408(83)90198-8
|
| [10] | von Alpen U, Bell MF, Höfer HH (1981) Compositional Dependence of the Electrochemical and Structural Parameters in the NASICON System (Na1+xSixZr2P3−xO12). Solid State Ionics 3–4: 215–218. |
| [11] |
Boilot JP, Salanié JP, Desplanches G, et al. (1979) Phase Transformation in Na1+xSixZr2P3–xO12 Compounds. Mater Res Bull 14: 1469–1477. doi: 10.1016/0025-5408(79)90091-6
|
| [12] |
Kohler H, Schultz H, Melnikov O (1983) Composition and Conduction Mechanism of the NASICON Structure-X-Ray Diffraction Study on two Crystals at Different Temperatures. Mater Res Bull 18: 1143–1152. doi: 10.1016/0025-5408(83)90158-7
|
| [13] |
Tsai CL, Hong HYP (1983) Investigation of phases and stability of solid electrolytes in the NASICON system. Mater Res Bull 18: 1399–1407. doi: 10.1016/0025-5408(83)90048-X
|
| [14] | Clearfield A, Subramanian MA, Wang W, et al. (1983) The Use of Hydrothermal Procedures to Synthesize NASICON and Some Comments on the Stoichiometry of NASICON Phases. Solid State Ionics 9–10: 895–902. |
| [15] |
Kohler H, Schultz H (1985) NASICON Solid Electrolytes Part I: The Na+-Diffusion Path and Its Relation to the Structure. Mater Res Bull 20: 1461–1471. doi: 10.1016/0025-5408(85)90164-3
|
| [16] |
Boilot JP, Collin G, Comes R (1983) Zirconium Deficiency in NASICON-Type Compounds: Crystal Structure of Na5Zr(PO4)3. J Solid State Chem 50: 91–99. doi: 10.1016/0022-4596(83)90236-0
|
| [17] | Boilot JP, Collin G, Comes R (1983) Stoichiometry and phase transitions in NASICON type compounds. Solid State Ionics 9–10: 829–834. |
| [18] |
Boilot JP, Collin G, Colomban Ph (1987) Crystal Structure of the True NASICON: Na3Zr2Si2PO12. Mater Res Bull 22: 669–676. doi: 10.1016/0025-5408(87)90116-4
|
| [19] | Boilot JP, Colomban Ph, Collin G (1988) Stoichiometry-Structure-Fast Ion Conduction in the NASICON Solid Solution. Solid State Ionics 28–30: 403–410. |
| [20] | Lucco-Borlera M, Mazza D, Montanaro L, et al. (1997) X-ray characterization of the new NASICON compositions Na3Zr2−x/4Si2−xP1+xO12 with x = 0.333, 0.667, 1.000, 1.333, 1.667. Powder Diffr 12: 171–174. |
| [21] |
Bohnke O, Ronchetti S, Mazza D (1999) Conductivity measurements on NASICON and nasicon-modified materials. Solid State Ionics 122: 127–136. doi: 10.1016/S0167-2738(99)00062-4
|
| [22] |
Naqash S, Ma Q, Tietz F, et al. (2017) Na3Zr2(SiO4)2(PO4) prepared by a solution-assisted solid state reaction. Solid State Ionics 302: 83–91. doi: 10.1016/j.ssi.2016.11.004
|
| [23] | Engell J, Mortensen S, Møller L (1983) Fabrication of NASICON Electrolytes from Metal Alkoxide Derived Gels. Solid State Ionics 9–10: 877–884. |
| [24] | Warhus U (1986) Synthese und Stabilität des NASICON Mischkristallsystems (Na1+xZr2SixP3−xO12, 0 ≤ x ≤ 3) [PhD thesis]. University of Stuttgart. |
| [25] | Kreuer KD, Kohler H, Maier J (1989) Sodium Ion Conductors with NASICON Framework Structure, In: Takahashi T, High Conductivity Ionic Conductors: Recent Trends and Application, Singapore: World Scientific Publishing Co., 242–279. |
| [26] |
Warhus U, Maier J, Rabenau A (1988) Thermodynamics of NASICON (Na1+xZr2SixP3−xO12). J Solid State Chem 72: 113–125. doi: 10.1016/0022-4596(88)90014-X
|
| [27] |
Mason TO, Hummel FA (1974) Compatibility Relations in the System SiO2-ZrO2-P2O5. J Am Ceram Soc 57: 538–539. doi: 10.1111/j.1151-2916.1974.tb10808.x
|
| [28] | Turkdogan ET, Maddocks WR (1952) Phase Equilibrium Investigation of the Na2O-SiO2-P2O5 Ternary System. J Iron Steel Inst 172: 1–15. |
| [29] | Milne SJ, West AR (1983) Compound formation and conductivity in the system Na2O-ZrO2-P2O5: Sodium zirconium orthophosphates. Solid State Ionics 9–10: 865–868. |
| [30] |
Milne SJ, West AR (1985) Zr-Doped Na3PO4: Crystal Chemistry, Phase Relations, and Polymorphism. J Solid State Chem 57: 166–177. doi: 10.1016/S0022-4596(85)80006-2
|
| [31] | Vlna M, Petrosyan Y, Kovar V, et al. (1993) Phase Coexistence in the System Na2O-P2O5-ZrO2. Chem Pap 47: 296–297. |
| [32] |
D'Ans J, Löffler J (1930) Untersuchungen im System Na2O-SiO2-ZrO2. Z Anorg Allg Chem 191: 1–35. doi: 10.1002/zaac.19301910102
|
| [33] | Polezhaev YM, Chukhlantsev VG (1965) Triangulation of the System Na2O-ZrO2-SiO2. Izv Akad Nauk SSSR Neorg Mater 1: 1990–1993; Inorg Mater (Engl Transl) 1: 1797–1800. |
| [34] | Sircar A, Brett NH (1970) Phase Equilibria in Ternary Systems Containing Zirconia and Silica, IV. Preliminary Study of the System Na2O-ZrO2-SiO2. Trans Brit Ceram Soc 69: 131–135. |
| [35] | Wilson G, Glasser FP (1987) Subsolidus Phase Relations in the System Na2O-ZrO2-SiO2. Brit Ceram Trans J 86: 199–201. |
| [36] |
Nicholas VA, Heyns AM, Kingon AI, et al. (1986) Reactions in the formation of Na3Zr2Si2PO12. J Mater Sci 21: 1967–1973. doi: 10.1007/BF00547935
|
| [37] |
Rudolf PR, Subramanian MA, Clearfield A, et al. (1985) The Crystal Structure of a Nonstoichiometric NASICON. Mater Res Bull 20: 643–651. doi: 10.1016/0025-5408(85)90142-4
|
| [38] |
Rudolf PR, Clearfield A, Jorgensen JD (1986) Rietveld Refinement Results on Three Nonstoichiometric Monoclinic NASICONs. Solid State Ionics 21: 213–224. doi: 10.1016/0167-2738(86)90075-5
|
| [39] |
Yde-Andersen S, Lundgaard JS, Møller L, et al. (1984) Properties of NASICON Electrolytes Prepared from Alkoxide Derived Gels: Ionic Conductivity, Durability in Molten Sodium and Strength Test Data. Solid State Ionics 14: 73–79. doi: 10.1016/0167-2738(84)90014-6
|
| [40] |
Perthuis H, Colomban Ph (1986) Sol-Gel Routes Leading to Nasicon Ceramics. Ceram Int 12: 39–52. doi: 10.1016/S0272-8842(86)80008-6
|
| [41] | Maier J, Warhus U, Gmelin E (1986) Thermodynamic and Electrochemical Investigations of the NASICON Solid Solution System. Solid State Ionics 18–19: 969–973. |
| [42] |
Boilot JP, Collin G, Colomban Ph (1988) Relation Structure-Fast Ion Conduction in the NASICON Solid Solution. J Solid State Chem 73: 160–171. doi: 10.1016/0022-4596(88)90065-5
|
| [43] |
Colomban Ph (1986) Orientational Disorder, Glass/Crystal Transition and Superionic Conductivity in NASICON. Solid State Ionics 21: 97–115. doi: 10.1016/0167-2738(86)90201-8
|
| [44] |
Park H, Jung K, Nezafati M, et al. (2016) Sodium Ion Diffusion in Nasicon (Na3Zr2Si2PO12) Solid Electrolytes: Effects of Excess Sodium. ACS Appl Mater Interfaces 8: 27814–27824. doi: 10.1021/acsami.6b09992
|
| [45] | Kafalas JA, Cava JR (1979) Effect of Pressure and Composition of Fast Na+-Ion Transport in the System Na1+xZr2SixP3−xO12. Proceedings of the International Conference on Fast Ion Transport in Solids, 419–422. |
| [46] |
Kreuer KD, Warhus U (1986) NASICON Solid Electrolytes: Part IV Chemical Durability. Mater Res Bull 21: 357–363. doi: 10.1016/0025-5408(86)90193-5
|
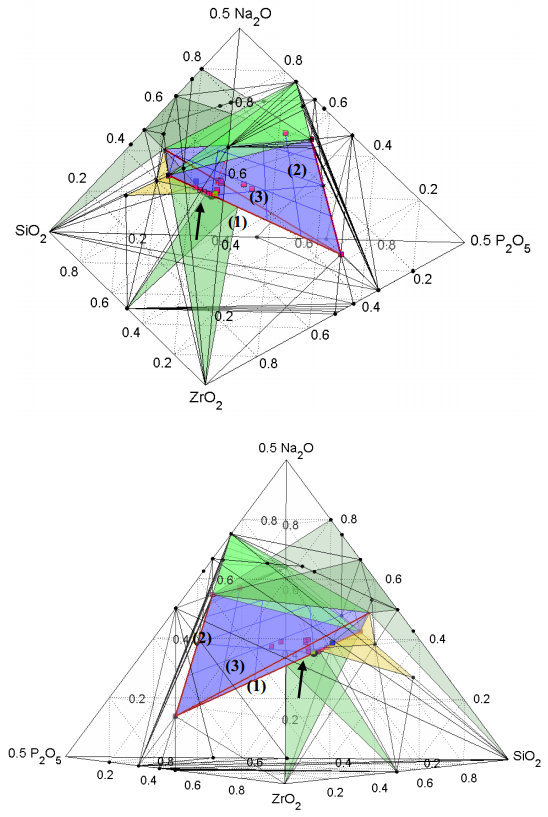
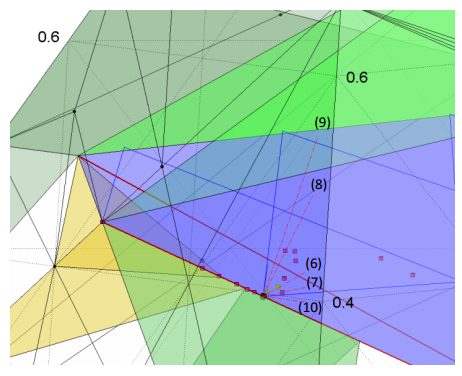

Figures(7) / Tables(1)

Frank Tietz. Phase relations of NASICON materials and compilation of the quaternary phase diagram Na2O-P2O5-SiO2-ZrO2[J]. AIMS Materials Science, 2017, 4(6): 1305-1318. doi: 10.3934/matersci.2017.6.1305







 DownLoad:
DownLoad: