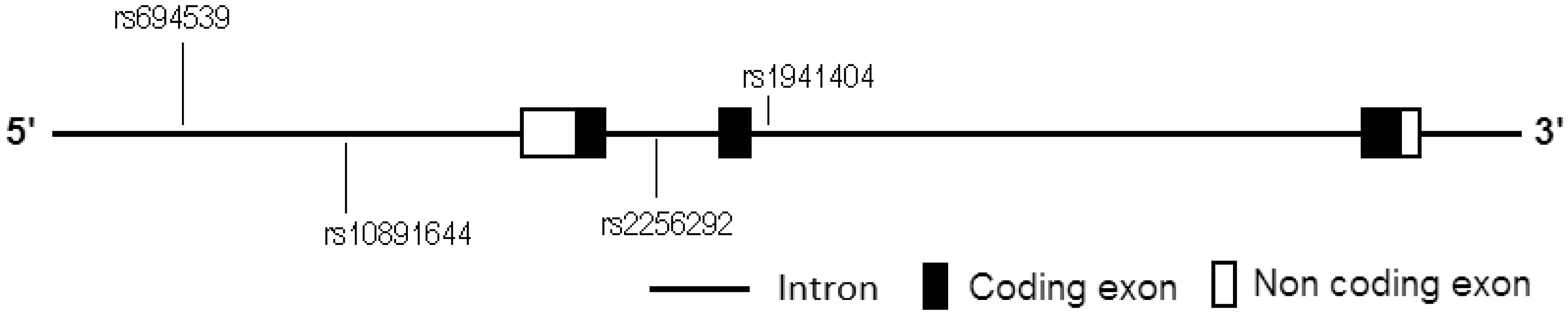
Although no direct evidence shows that Nicotinamide N-methyltransferase (NNMT) is involved in hypertension to date, many reports have shown that NNMT plays important roles in pathogenesis of metabolic syndrome and in various cardiovascular diseases. Here we explored the genetic association between NNMT polymorphisms and primary hypertension. 308 primary hypertension cases (aged < 60 years, blood pressure ≥ 140/90 mmHg or taking antihypertensive therapies) and 315 controls (aged ≥ 60 years and without any diagnosed diseases) were recruited from unrelated Chinese Han ethnicity volunteers. Then a case-control study was carried out to explore the genetic association between NNMT polymorphism and primary hypertension. A significantly associated SNP (rs1941404) was found in NNMT gene (P < 0.0125). At this locus, the risks for minor homozygote CC carriers being of hypertension were highly significantly higher than those for the TT + CT carriers (OR = 2.120, ORadjusted = 2.573, P = 0.000, Padjusted = 0.001 and statistical power = 0.997), but the differences between the genotypes CT + CC and TT (dominant genetic model) were not statistically significant (P = 0.321, Padjusted = 0.230 and statistical power = 0.200). In conclusion, rs1941404 in NNMT gene sequence is significantly associated with primary hypertension under a recessive inheritance mode. At this locus, the minor homozygote CC carriers are susceptible population and the TT and CT carriers are non-susceptible population.
Citation: Xiang-Xiang Guan, Xiao-Juan Zhu, Zhao-Hui Deng, Yu-Rong Zeng, Jie-Ru Liu, Jiang-Hua Li. The association between nicotinamide N-methyltransferase gene polymorphisms and primary hypertension in Chinese Han Population[J]. AIMS Bioengineering, 2021, 8(2): 130-139. doi: 10.3934/bioeng.2021012
Although no direct evidence shows that Nicotinamide N-methyltransferase (NNMT) is involved in hypertension to date, many reports have shown that NNMT plays important roles in pathogenesis of metabolic syndrome and in various cardiovascular diseases. Here we explored the genetic association between NNMT polymorphisms and primary hypertension. 308 primary hypertension cases (aged < 60 years, blood pressure ≥ 140/90 mmHg or taking antihypertensive therapies) and 315 controls (aged ≥ 60 years and without any diagnosed diseases) were recruited from unrelated Chinese Han ethnicity volunteers. Then a case-control study was carried out to explore the genetic association between NNMT polymorphism and primary hypertension. A significantly associated SNP (rs1941404) was found in NNMT gene (P < 0.0125). At this locus, the risks for minor homozygote CC carriers being of hypertension were highly significantly higher than those for the TT + CT carriers (OR = 2.120, ORadjusted = 2.573, P = 0.000, Padjusted = 0.001 and statistical power = 0.997), but the differences between the genotypes CT + CC and TT (dominant genetic model) were not statistically significant (P = 0.321, Padjusted = 0.230 and statistical power = 0.200). In conclusion, rs1941404 in NNMT gene sequence is significantly associated with primary hypertension under a recessive inheritance mode. At this locus, the minor homozygote CC carriers are susceptible population and the TT and CT carriers are non-susceptible population.

| [1] |
Pissios P (2017) Nicotinamide N-methyltransferase: more than a vitamin B3 clearance enzyme. Trends Endocrinol Metab 28: 340-353. doi: 10.1016/j.tem.2017.02.004

|
| [2] |
Li JH, Qiu LQ, Zhu XJ, et al. (2017) Influence of exercises using different energy metabolism systems on NNMT expression in different types of skeletal muscle fibers. Sci Sport 32: 27-32. doi: 10.1016/j.scispo.2016.06.004
|
| [3] |
Li F, Chong ZZ, Maiese K (2006) Cell Life versus cell longevity: the mysteries surrounding the NAD+ precursor nicotinamide. Curr Med Chem 13: 883-895. doi: 10.2174/092986706776361058
|
| [4] |
Bromberg A, Levine J, Belmaker R, et al. (2011) Hyperhomocysteinemia does not affect global DNA methylation and nicotinamide N-methyltransferase expression in mice. J Psychopharmacol 25: 976-981. doi: 10.1177/0269881110388328
|
| [5] |
Souto JC, Blanco-Vaca F, Soria JM, et al. (2005) A genomewide exploration suggests a new candidate gene atchromosome 11q23 as themajorde terminant of plasma homocysteine levels: results from the GAIT project. Am J Hum Genet 76: 925-933. doi: 10.1086/430409
|
| [6] |
Williams AC, Hill LJ, Ramsden DB (2012) Nicotinamide, NAD (P)(H), and methyl-group homeostasis evolved and became a determinant of ageing diseases: hypotheses and lessons from pellagra. Curr Gerontol Geriatr Res 2012: 302875. doi: 10.1155/2012/302875
|
| [7] |
Giuliante R, Sartini D, Bacchetti T, et al. (2015) Potential involvement of nicotinamide N-methyltransferase in the pathogenesis of metabolic syndrome. Metab Syndr Relat Disord 13: 165-170. doi: 10.1089/met.2014.0134
|
| [8] |
Li JH, Wang YH, Zhu XJ, et al. (2018) Metabolomics study on the association between nicotinamide N-methyltransferase gene polymorphisms and type 2 diabetes. Int J Diabetes Dev Ctries 38: 409-416. doi: 10.1007/s13410-017-0601-2
|
| [9] |
Giusti B, Saracini C, Bolli P, et al. (2008) Genetic analysis of 56 polymorphisms in 17 genes involved in methionine metabolism in patients with abdominal aortic aneurysm. J Med Genet 45: 721-730. doi: 10.1136/jmg.2008.057851
|
| [10] |
Sazci A, Sazci G, Sazci B, et al. (2016) Nicotinamide-N-Methyltransferase gene rs694539 variant and migraine risk. J Headache Pain 17: 93. doi: 10.1186/s10194-016-0688-8
|
| [11] |
van Driel LM, Smedts HP, Helbing WA, et al. (2008) Eight-fold increased risk for congenital heart defects in children carrying the nicotinamide N-methyltransferase polymorphism and exposed to medicines and low nicotinamide. Eur Heart J 29: 1424-1431. doi: 10.1093/eurheartj/ehn170
|
| [12] |
Sazci A, Ozel MD, Ergul E, et al. (2013) Association of nicotinamide-N-methyltransferase (NNMT) gene rs694539 variant with bipolar disorder. Gene 532: 272-275. doi: 10.1016/j.gene.2013.08.077
|
| [13] |
Sazci A, Ozel MD, Ergul E, et al. (2013) Association of nicotinamide-N-methyltransferase gene rs694539 variant with patients with nonalcoholic steatohepatitis. Genet Test Mol Biomarkers 17: 849-853. doi: 10.1089/gtmb.2013.0309
|
| [14] |
Sazci G, Sazci B, Sazci A, et al. (2016) Association of Nicotinamide-N-Methyltransferase Gene rs694539 Variant with Epilepsy. Mol Neurobiol 53: 4197-4200. doi: 10.1007/s12035-015-9364-2
|
| [15] | Bromberg A, Lerer E, Udawela M, et al. (2012) Nicotinamide-N-methyltransferase (NNMT) in schizophrenia: genetic association and decreased frontal cortex mRNA levels. Int J Neuro psycho pharmacol 15: 727-737. |
| [16] | Zhou Q, Zhu XJ, Li JH (2017) Association between nicotinamide N-methyltransferase gene polymorphisms and obesity in Chinese Han Male College Students. Biomed Res Int 2017: 2984826. |
| [17] | Zhu XJ, Lin YJ, Chen W, et al. (2016) Physiological study on association between nicotinamide N-methyltransferase gene polymorphisms and hyperlipidemia. Biomed Res Int 2016: 7521942. |
| [18] |
Li JH, Chen W, Zhu XJ, et al. (2017) Associations of nicotinamide N-methyltransferase gene single nucleotide polymorphisms with sport performance and relative maximal oxygen uptake. J Sport Sci 35: 2185-2190. doi: 10.1080/02640414.2016.1261176
|
| [19] | Azam AB, Azizan EAB (2018) Brief overview of a decade of genome-wide association studies on primary hypertension. Int J Endocrinol 2018: 7259704. |
| [20] |
Shi YY, He L (2005) SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res 15: 97-98. doi: 10.1038/sj.cr.7290272
|
| [21] |
Oparil S, Zaman MA, Calhoun DA (2003) Pathogenesis of hypertension. Ann Intern Med 139: 761-776. doi: 10.7326/0003-4819-139-9-200311040-00011
|
| [22] |
Pavlou DI, Paschou SA, Anagnostis P, et al. (2018) Hypertension in patients with type 2 diabetes mellitus: Targets and management. Maturitas 112: 71-77. doi: 10.1016/j.maturitas.2018.03.013
|
| [23] |
Lastra G, Syed S, Kurukulasuriya LR, et al. (2014) Type 2 diabetes mellitus and hypertension: an update. Endocrinol Metab Clin North Am 43: 103-122. doi: 10.1016/j.ecl.2013.09.005
|
| [24] |
Lima NK, Abbasi F, Lamendola C, et al. (2009) Prevalence of insulin resistance and related risk factors for cardiovascular disease in patients with essential hypertension. Am J Hypertens 22: 106-111. doi: 10.1038/ajh.2008.263
|
| [25] |
Bonora E, Capaldo B, Perin PC, et al. (2008) Hyperinsulinemia and insulin resistance are independently associated with plasma lipids, uric acid and blood pressure in non-diabetic subjects. The GISIR database. Nutr Metab Cardiovasc Dis 18: 624-631. doi: 10.1016/j.numecd.2007.05.002
|
| [26] | Shi L, Liu XY, Huang ZG, et al. (2019) Endogenous hydrogen sulfide and ERK1/2-STAT3 signaling pathway may participate in the association between homocysteine and hypertension. J Geriatr Cardiol 16: 822-834. |
| [27] |
Hidru TH, Yang X, Xia Y, et al. (2019) The relationship between plasma markers and essential hypertension in middle-aged and elderly Chinese population: a community based cross-sectional study. Sci Rep 9: 6813. doi: 10.1038/s41598-019-43278-4
|
| [28] |
Zhang L, Miyaki K, Araki J, et al. (2007) The relation between nicotinamide N-methyltransferase gene polymorphism and plasma homocysteine concentration in healthy Japanese men. Thromb Res 121: 55-58. doi: 10.1016/j.thromres.2007.02.012
|
| [29] |
Ramsden DB, Waring RH, Parsons RB, et al. Nicotinamide N-methyltransferase: genomic connection to disease (2020) . doi: 10.1177/1178646920919770
|
| [30] |
Yan L, Otterness DM, Weinshilboum RM (1999) Human nicotinamide N-methyltransferase pharmacogenetics: gene sequence analysis and promoter characterization. Pharmacogenetics 9: 307-316. doi: 10.1097/00008571-199906000-00005
|
| [31] |
Smith ML, Burnett D, Bennett P, et al. (1998) A direct correlation between nicotinamide N-methyltransferase activity and protein levels in human liver cytosol. Biochim Biophys Acta 1442: 238-244. doi: 10.1016/S0167-4781(98)00177-8
|
Figures(1) / Tables(5)

Xiang-Xiang Guan, Xiao-Juan Zhu, Zhao-Hui Deng, Yu-Rong Zeng, Jie-Ru Liu, Jiang-Hua Li. The association between nicotinamide N-methyltransferase gene polymorphisms and primary hypertension in Chinese Han Population[J]. AIMS Bioengineering, 2021, 8(2): 130-139. doi: 10.3934/bioeng.2021012







 DownLoad:
DownLoad: