Citation: I. Banegas, I. Prieto, A.B. Segarra, M. Martínez-Cañamero, M. de Gasparo, M. Ramírez-Sánchez. Angiotensin II, dopamine and nitric oxide. An asymmetrical neurovisceral interaction between brain and plasma to regulate blood pressure[J]. AIMS Neuroscience, 2019, 6(3): 116-127. doi: 10.3934/Neuroscience.2019.3.116

| [1] |
Oparil S, Acelajado MC, Bakris GL, et al. (2018) Hypertension. Nat Rev Dis Primers 4: 18014. doi: 10.1038/nrdp.2018.14

|
| [2] |
Prieto I, Villarejo AB, Segarra AB, et al. (2014) Brain, heart and kidney correlate for the control of blood pressure and water balance: role of angiotensinases. Neuroendocrinology 100: 198–208. doi: 10.1159/000368835
|
| [3] |
Prieto I, Segarra AB, Martinez-Canamero M, et al. (2017) Bidirectional asymmetry in the neurovisceral communication for the cardiovascular control: New insights. Endocr Regul 51: 157–167. doi: 10.1515/enr-2017-0017
|
| [4] | Prieto I, Segarra AB, de Gasparo M, et al. (2018) Divergent profile between hypothalamic and plasmatic aminopeptidase activities in WKY and SHR. Influence of beta-adrenergic blockade. Life Sci 192: 9–17. |
| [5] |
Parati G, Ochoa JE, Lombardi C, et al. (2013) Assessment and management of blood-pressure variability. Nat Rev Cardiol 10: 143–155. doi: 10.1038/nrcardio.2013.1
|
| [6] |
Banegas I, Prieto I, Vives F, et al. (2006) Brain aminopeptidases and hypertension. J Renin Angiotensin Aldosterone Syst 7: 129–134. doi: 10.3317/jraas.2006.021
|
| [7] |
Banegas I, Prieto I, Vives F, et al. (2009) Asymmetrical response of aminopeptidase A and nitric oxide in plasma of normotensive and hypertensive rats with experimental hemiparkinsonism. Neuropharmacology 56: 573–579. doi: 10.1016/j.neuropharm.2008.10.011
|
| [8] |
Banegas I, Prieto I, Segarra AB, et al. (2011) Blood pressure increased dramatically in hypertensive rats after left hemisphere lesions with 6-hydroxydopamine. Neurosci Lett 500: 148–150. doi: 10.1016/j.neulet.2011.06.025
|
| [9] |
Banegas I, Prieto I, Segarra AB, et al. (2017) Bilateral distribution of enkephalinase activity in the medial prefrontal cortex differs between WKY and SHR rats unilaterally lesioned with 6-hydroxydopamine. Prog Neuropsychopharmacol Biol Psychiatry 75: 213–218. doi: 10.1016/j.pnpbp.2017.02.015
|
| [10] |
Prieto I, Segarra AB, Villarejo AB, et al. (2019) Neuropeptidase activity in the frontal cortex of Wistar-Kyoto and spontaneously hypertensive rats treated with vasoactive drugs: a bilateral study. J Hypertens 37: 612–628. doi: 10.1097/HJH.0000000000001884
|
| [11] | Cuspidi C, Tadic M, Grassi G, et al. (2018) Mancia G. Treatment of hypertension: The ESH/ESC guidelines recommendations. Pharmacol Res 128: 315–321. |
| [12] |
Ramírez-Sánchez M, Prieto I, Wangensteen R, et al. (2013) The renin-angiotensin system: new insight into old therapies. Curr Med Chem 20: 1313–1322. doi: 10.2174/0929867311320100008
|
| [13] |
Ferdinand KC, Balavoine F, Besse B, et al. (2019) Efficacy and safety of Firibastat, a first-in-class brain aminopeptidase A inhibitor, in hypertensive overweight patients of multiple ethnic origins: A Phase 2, open-label, multicenter, dose-titrating study. Circulation 140: 138–146. doi: 10.1161/CIRCULATIONAHA.119.040070
|
| [14] |
Gao J, Marc Y, Iturrioz X, et al. (2014) A new strategy for treating hypertension by blocking the activity of the brain renin-angiotensin system with aminopeptidase A inhibitors. Clin Sci (Lond) 127: 135–148. doi: 10.1042/CS20130396
|
| [15] |
Keck M, De Almeida H, Compère D, et al. (2019) NI956/QGC006, a potent orally active, brain-penetrating aminopeptidase a inhibitor for treating hypertension. Hypertension 73: 1300–1307. doi: 10.1161/HYPERTENSIONAHA.118.12499
|
| [16] |
Wright JW, Mizutani S, Murray CE, et al. (1990) Aminopeptidase-induced elevations and reductions in blood pressure in the spontaneously hypertensive rat. J Hypertens 8: 969–974. doi: 10.1097/00004872-199010000-00013
|
| [17] |
Wright JW, Jensen LL, Cushing LL, et al. (1989) Leucine aminopeptidase M-induced reductions in blood pressure in spontaneously hypertensive rats. Hypertension 13: 910–915. doi: 10.1161/01.HYP.13.6.910
|
| [18] |
Zini S, Masdehors P, Lenkei Z, et al. (1997) Aminopeptidase A: distribution in rat brain nuclei and increased activity in spontaneously hypertensive rats. Neuroscience 78: 1187–1193. doi: 10.1016/S0306-4522(96)00660-4
|
| [19] |
Banegas I, Prieto I, Segarra AB, et al. (2017) Study of the neuropeptide function in Parkinson's disease using the 6-Hydroxydopamine model of experimental Hemiparkinsonism. AIMS Neuroscience 4: 223–237. doi: 10.3934/Neuroscience.2017.4.223
|
| [20] |
Kondo K, Ebihara A, Suzuki H, et al. (1981) Role of dopamine in the regulation of blood pressure and the renin--angiotensin-aldosterone system in conscious rats. Clin Sci (Lond) 61: 235s–237s. doi: 10.1042/cs061235s
|
| [21] |
Zeng C, Jose PA (2011) Dopamine receptors: important antihypertensive counterbalance against hypertensive factors. Hypertension 57: 11–17. doi: 10.1161/HYPERTENSIONAHA.110.157727
|
| [22] |
Förstermann U, Sessa WC (2012) Nitric oxide synthases: regulation and function. Eur Heart J 33: 829–837. doi: 10.1093/eurheartj/ehr304
|
| [23] | Calver A, Collier J, Vallance P (1993) Nitric oxide and the control of human vascular tone in health and disease. Eur J Med 2: 48–53. |
| [24] |
Venturelli M, Pedrinolla A, Boscolo Galazzo I, et al. (2018) Impact of nitric oxide bioavailability on the progressive cerebral and peripheral circulatory impairments during aging and Alzheimer's disease. Front Physiol 9: 169. doi: 10.3389/fphys.2018.00169
|
| [25] |
Steinert JR, Chernova T, Forsythe ID (2010) Nitric oxide signaling in brain function, dysfunction, and dementia. Neuroscientist 16: 435–452. doi: 10.1177/1073858410366481
|
| [26] | Maia-de-Oliveira JP, Trzesniak C, Oliveira IR, et al. (2012) Nitric oxide plasma/serum levels in patients with schizophrenia: a systematic review and meta-analysis. Braz J Psychiatry 34: S149–155. |
| [27] |
Nakano Y, Yoshimura R, Nakano H, et al. (2010) Association between plasma nitric oxide metabolites levels and negative symptoms of schizophrenia: a pilot study. Hum Psychopharmacol 25: 139–144. doi: 10.1002/hup.1102
|
| [28] |
Shabeeh H, Khan S, Jiang B, et al. (2017) Blood pressure in healthy humans is regulated by neuronal no synthase. Hypertension 69: 970–976. doi: 10.1161/HYPERTENSIONAHA.116.08792
|
| [29] |
Chen ZQ, Mou RT, Feng DX, et al. (2017) The role of nitric oxide in stroke. Med Gas Res 7: 194–203. doi: 10.4103/2045-9912.215750
|
| [30] |
Zheng R, Qin L, Li S, et al. (2014) CT perfusion-derived mean transit time of cortical brain has a negative correlation with the plasma level of nitric oxide after subarachnoid hemorrhage. Acta Neurochir (Wien) 156: 527–533. doi: 10.1007/s00701-013-1968-6
|
| [31] |
Taffi R, Nanetti L, Mazzanti L, et al. (2008) Plasma levels of nitric oxide and stroke outcome. J Neurol 255: 94–98. doi: 10.1007/s00415-007-0700-y
|
| [32] |
Li S, Wang Y, Jiang Z, et al. (2018) Impaired cognitive performance in endothelial nitric oxide synthase knockout mice after ischemic stroke: A pilot study. Am J Phys Med Rehabil 97: 492–499. doi: 10.1097/PHM.0000000000000904
|
| [33] | Bernard C (1878) Etude sur la physiologie du coeur. La science experimentale: 316–366. Available from: https://gallica.bnf.fr/ark:/12148/bpt6k69992b/f316.image. |
| [34] |
Thayer JF, Lane RD (2009) Claude Bernard and the heart-brain connection: further elaboration of a model of neurovisceral integration. Neurosci Biobehav Rev 33: 81–88. doi: 10.1016/j.neubiorev.2008.08.004
|
| [35] |
Segarra AB, Prieto I, Banegas I, et al. (2012) Asymmetrical effect of captopril on the angiotensinase activity in frontal cortex and plasma of the spontaneously hypertensive rats: expanding the model of neuroendocrine integration. Behav Brain Res 230: 423–427. doi: 10.1016/j.bbr.2012.02.039
|
| [36] | Segarra AB, Prieto I, Banegas I, et al. (2013) The brain-heart connection: frontal cortex and left ventricle angiotensinase activities in control and captopril-treated hypertensive rats-a bilateral study. Int J Hypertens 2013: 156179. |
| [37] | Segarra AB, Banegas I, Prieto I, et al. (2016) Brain asymmetry and dopamine: beyond motor implications in Parkinson's disease and experimental hemiparkinsonism. Rev Neurol 63: 415–421. |
| [38] |
de Jager L, Amorim EDT, Lucchetti BFC, et al. (2018) Nitric oxide alterations in cardiovascular system of rats with Parkinsonism induced by 6-OHDA and submitted to previous exercise. Life Sci 204: 78–86. doi: 10.1016/j.lfs.2018.05.017
|
| [39] |
Alexander N, Kaneda N, Ishii A, et al. (1990) Right-left asymmetry of tyrosine hydroxylase in rat median eminence: influence of arterial baroreflex nerves. Brain Res 523: 195–198. doi: 10.1016/0006-8993(90)91487-2
|
| [40] |
Hersh LB (1985) Characterization of membrane-bound aminopeptidases from rat brain:identification of the enkephalin-degrading aminopeptidase. J Neurochem 44: 1427–1435. doi: 10.1111/j.1471-4159.1985.tb08779.x
|
| [41] |
Wright JW, Harding JW (1997) Important roles for angiotensin III and IV in the brain renin-angiotensin system. Brain Res Rev 25: 96–124. doi: 10.1016/S0165-0173(97)00019-2
|
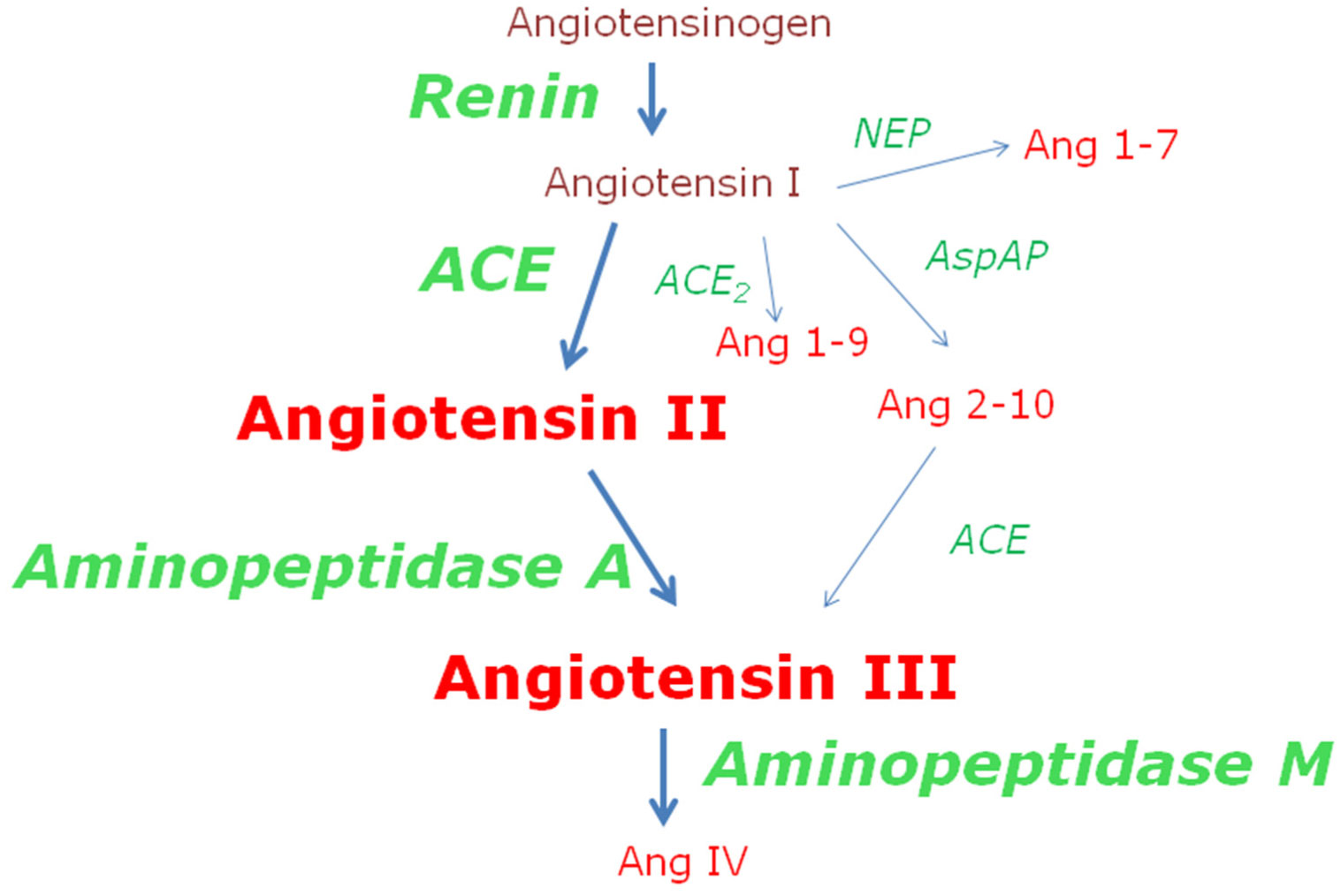
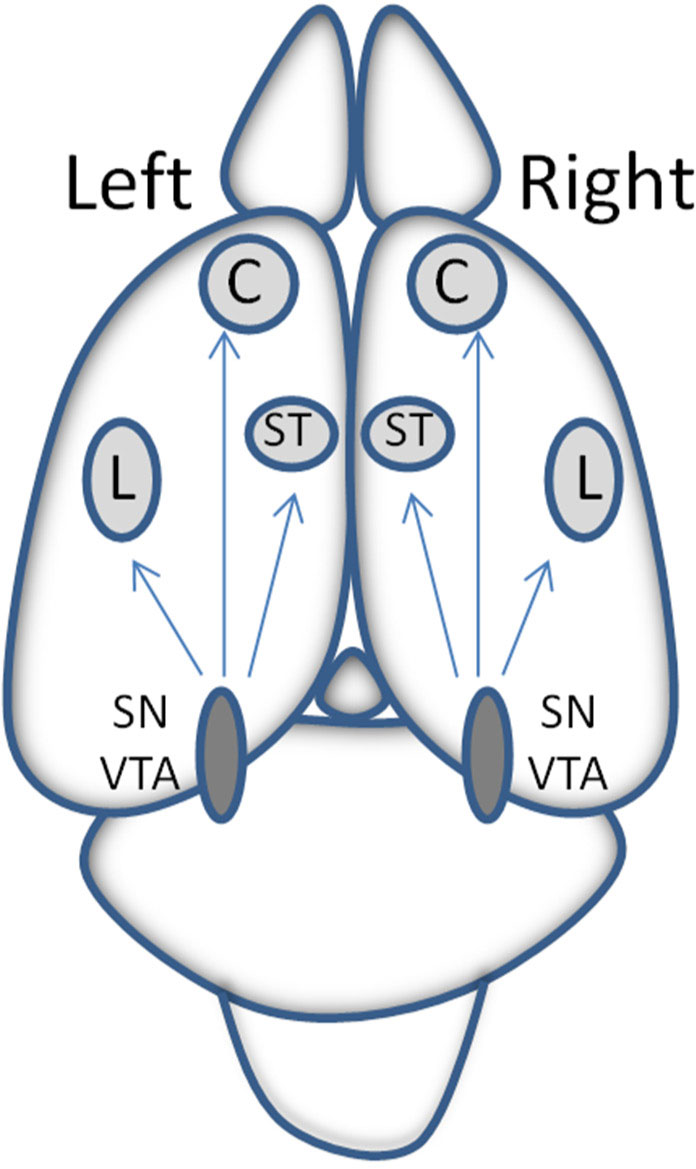
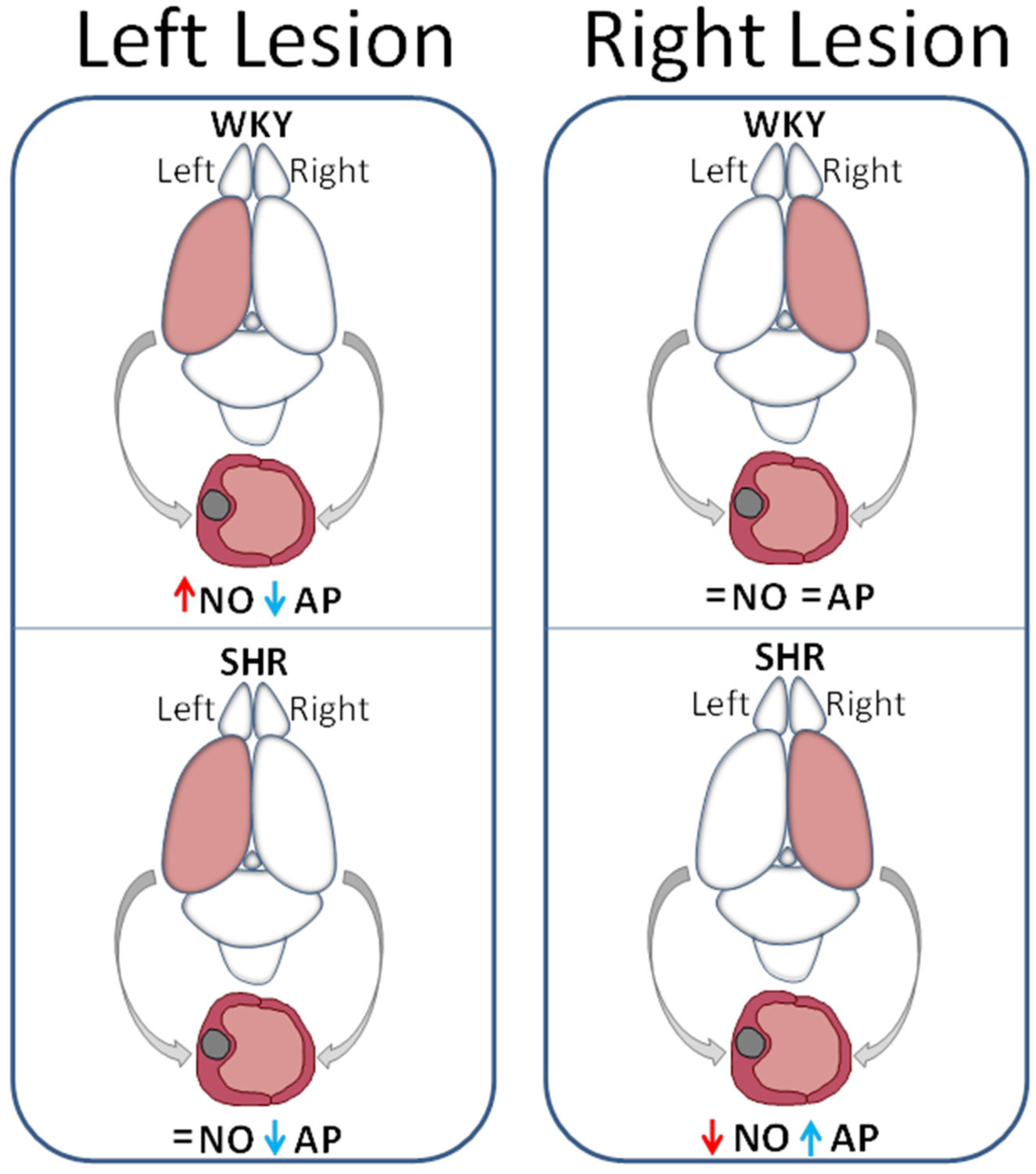
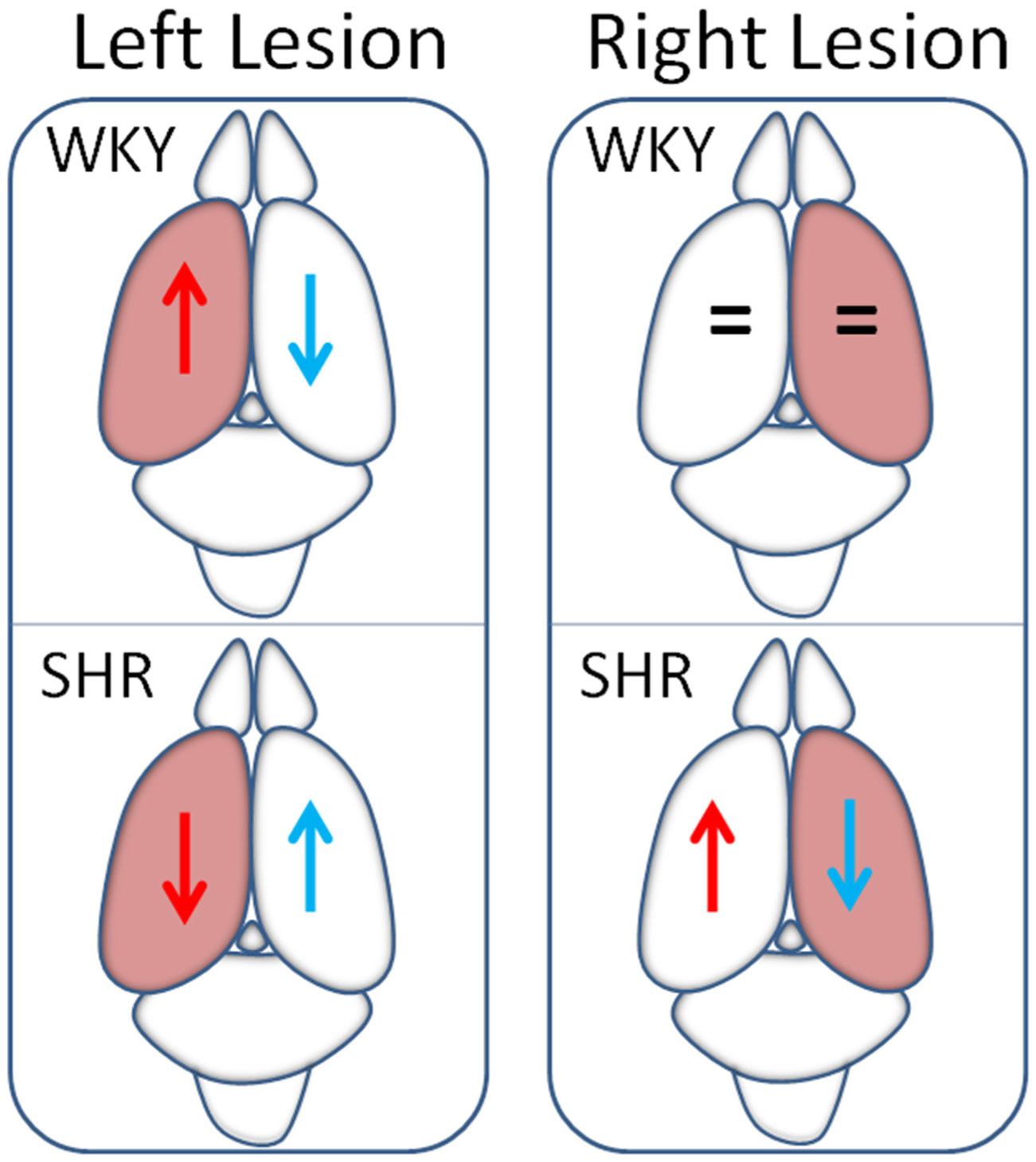
Figures(4)

I. Banegas, I. Prieto, A.B. Segarra, M. Martínez-Cañamero, M. de Gasparo, M. Ramírez-Sánchez. Angiotensin II, dopamine and nitric oxide. An asymmetrical neurovisceral interaction between brain and plasma to regulate blood pressure[J]. AIMS Neuroscience, 2019, 6(3): 116-127. doi: 10.3934/Neuroscience.2019.3.116







 DownLoad:
DownLoad: