Citation: Edward N Trifonov. Columnar structure of SV40 minichromosome[J]. AIMS Biophysics, 2015, 2(3): 274-283. doi: 10.3934/biophy.2015.3.274

| [1] |
Griffith JD (1975) Chromatin structure: deduced from a minichromosome. Science 187: 1202-1203. doi: 10.1126/science.187.4182.1202

|
| [2] |
Bellard M, Oudet P, Germond JE, et al. (1976) Subunit structure of simian-virus-40 minichromosome. Eur J Biochem 70: 543-553. doi: 10.1111/j.1432-1033.1976.tb11046.x
|
| [3] |
Mengeritsky G, Trifonov EN (1984) Nucleotide sequence-directed mapping of the nucleosomes of SV40 chromatin. Cell Bioph 6: 1-9. doi: 10.1007/BF02788576
|
| [4] |
Ambrose C, Lowman H, Rajadhyaksha A, et al. (1990) Location of nucleosomes in simian virus 40 chromatin. J Mol Biol 214: 875-884. doi: 10.1016/0022-2836(90)90342-J
|
| [5] | Tatchell K, van Holde KE (1978) Compact oligomers and nucleosome phasing. Proc Natl Acad Sci USA 75: 3583-3587. |
| [6] |
Fajkus J, Trifonov EN (2001) Columnar packing of telomeric nucleosomes. Biochem Biophys Res Comm 280: 961-963. doi: 10.1006/bbrc.2000.4208
|
| [7] | Salih B, Trifonov EN (2015) Strong nucleosomes of A. thaliana concentrate in centromere regions. J Biomol Str Dyn 33: 10-13. |
| [8] |
Salih B, Trifonov EN (2015). Strong nucleosomes reside in meiotic centromeres of C. elegans. J Biomol Str Dyn 33: 365-373. doi: 10.1080/07391102.2013.879263
|
| [9] |
Nibhani R, Trifonov EN (2015) Reading sequence-directed computational nucleosome maps. J Biomol Str Dyn 33:1558-1566. doi: 10.1080/07391102.2014.958877
|
| [10] |
Gu SG, Goszczynski B, McGhee JD, et al. (2013) Unusual DNA packaging characteristics in endoreduplicated Caenorhabditis elegans oocytes defined by in vivo accessibility to an endogenous nuclease activity. Epigenet Chromatin 6: 37. doi: 10.1186/1756-8935-6-37
|
| [11] |
Ponder BAJ, Crawford LV (1977) Arrangement of nucleosomes in nucleoprotein complexes from polyoma-virus and SV40. Cell 11: 35-49. doi: 10.1016/0092-8674(77)90315-4
|
| [12] | Zhang T, Talbert PB, Zhang W, et al. (2013) The CentO satellite confers translational and rotational phasing on cenH3 nucleosomes in rice centromeres. Proc Natl Acad Sci U S A 110: E4875-4883. |
| [13] | Rattner JB, Hamkalo BA (1978) Higher-order structure in metaphase chromosomes Chromosoma 69: 373-379. |
| [14] | Dubochet J, Adrian M, Schultz P, et al. (1986) Cryo-electron microscopy of vitrified SV40 minichromosomes: the liquid drop model. EMBO J 5: 519-528. |
| [15] |
Wanner G, Formanek H (2000) A new chromosome model. J Struct Biol 132: 147-161. doi: 10.1006/jsbi.2000.4310
|
| [16] |
Karpov VL, Bavykin SG, Preobrazhenskaya OV, et al. (1982) Alignment of nucleosomes along DNA and organization of spacer DNA in drosophila chromatin. Nucl Acids Res 10: 4321-4337. doi: 10.1093/nar/10.14.4321
|
| [17] | Widom J (1992) A relationship between the helical twist of DNA and the ordered positioning of nucleosomes in all eukaryotic cells. Proc Natl Acad Sci U S A 89:1095-1099. |
| [18] |
Rapoport AE, Frenkel ZM, Trifonov EN (2011) Nucleosome positioning pattern derived from oligonucleotide compositions of genomic sequences. J Biomol Str Dyn 28: 567-574. doi: 10.1080/07391102.2011.10531243
|
| [19] |
Frenkel ZM, Bettecken T, Trifonov EN (2011) Nucleosome DNA sequence structure of isochores. BMC Genomics 12: 203. doi: 10.1186/1471-2164-12-203
|
| [20] |
Tripathi V, Salih B, Trifonov EN (2015) Universal full-length nucleosome mapping sequence probe. J Biomol Str Dyn 33: 666-673. doi: 10.1080/07391102.2014.891262
|
| [21] |
Lowary PT, Widom J (1998) New DNA sequence rules for high affinity binding to histone octamer and sequence directed nucleosome positioning. J Mol Biol 276: 19-42. doi: 10.1006/jmbi.1997.1494
|
| [22] |
Trifonov EN, Nibhani R (2015) Review fifteen years of search for strong nucleosomes. Biopolymers, 103: 432-437. doi: 10.1002/bip.22604
|
| [23] | Nibhani R, Trifonov EN, TA-periodic (“601”-like) centromeric nucleosomes of A.thaliana. J Biomol Str Dyn [in press]. |
| [24] |
Vasudevan D, Chua EYD, Davey CA (2010) Crystal structures of nucleosome core particles containing the ‘601’ strong positioning sequence. J Mol Biol 403: 1-10. doi: 10.1016/j.jmb.2010.08.039
|
| [25] | Zhurkin VB (1982) Periodicity in DNA primary structure and specific alignment of nucleosomes. Stud Biophys 87: 151-152. |
| [26] |
Zhurkin VB (1983) Specific alignment of nucleosomes on DNA correlates with periodic distribution of purine pyrimidine and pyrimidine purine dimers. FEBS Lett 158: 293-297. doi: 10.1016/0014-5793(83)80598-5
|
| [27] |
Zhurkin VB, Lysov YP, Ivanov VI (1979) Anisotropic flexibility of DNA and the nucleosomal structure. Nucl Acids Res 6: 1081-1096. doi: 10.1093/nar/6.3.1081
|
| [28] |
Wang D, Ulyanov NB, Zhurkin VB (2010) Sequence-dependent kink-and-slide deformations of nucleosomal DNA facilitated by histone arginines bound in the minor groove. J Biomol Str Dyn 27: 843-859. doi: 10.1080/07391102.2010.10508586
|
| [29] | McDowall AW, Smith JM, Dubochet J (1986) Cryo-electron microscopy of vitrified chromosomes in situ. EMBO J 5: 1395-1402. |
| [30] | Trifonov EN Nucleosome repeat lengths and columnar chromatin structure. J Biomol Str Dyn [in press]. |
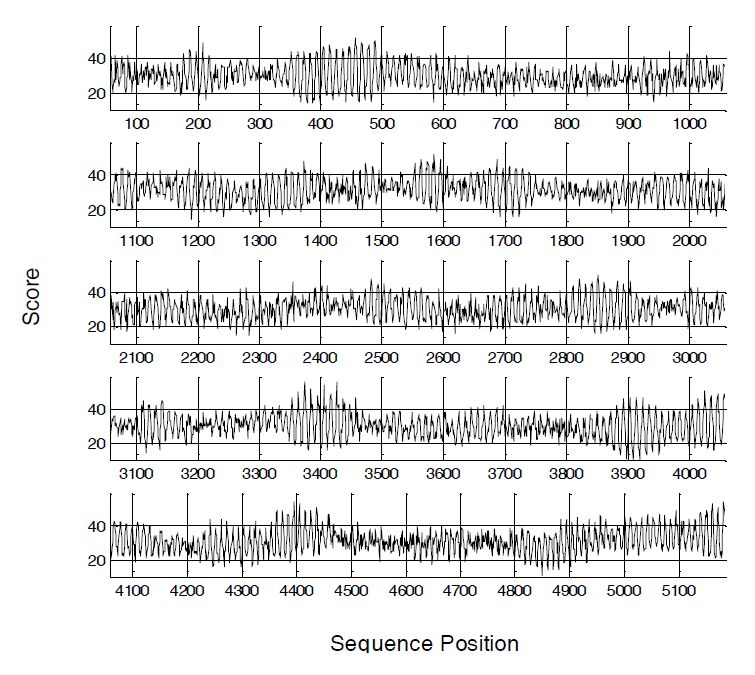

Figures(2) / Tables(1)

Edward N Trifonov. Columnar structure of SV40 minichromosome[J]. AIMS Biophysics, 2015, 2(3): 274-283. doi: 10.3934/biophy.2015.3.274







 DownLoad:
DownLoad: