Cancer represents a major global health burden, prompting continuous research for effective therapeutic strategies. Natural compounds derived from plants have emerged as potential strategies for preventing cancer and treatment because of their inherent pharmacological properties. This comprehensive review aimed to evaluate the therapeutic potential of five key natural compounds: apigenin, quercetin, piperine, curcumin, and resveratrol in cancer prevention and therapy. By examining their molecular mechanisms and preclinical evidence, this review seeks to elucidate their role as potential adjuvants or stand-alone therapies in cancer management. The exploration of natural compounds as cancer therapeutics offers several advantages, including low toxicity, wide availability, and compatibility with conventional chemotherapeutic agents. We highlighted the current understanding of their anticancer mechanisms and clinical applications for advancing personalized cancer care to improve patient outcomes. We discussed the empirical findings from in vitro, in vivo, and clinical studies reporting biological activity and therapeutic efficacy in antioxidant, immunomodulatory, anti-carcinogenic, and chemo-sensitizing modes. Innovative delivery systems and personalized treatment approaches may further enhance their bioavailability and therapeutic utility in a synergistic approach with chemo- and radiotherapeutic disease management. This review underscores the importance of natural compounds in cancer prevention and treatment, promoting a multidisciplinary approach to the development of innovative therapeutic strategies.
Citation: Amogh Auti, Madhura Tathode, Maria Michela Marino, Antonio Vitiello, Andrea Ballini, Francesco Miele, Valeria Mazzone, Alessia Ambrosino, Mariarosaria Boccellino. Nature's weapons: Bioactive compounds as anti-cancer agents[J]. AIMS Public Health, 2024, 11(3): 747-772. doi: 10.3934/publichealth.2024038
Cancer represents a major global health burden, prompting continuous research for effective therapeutic strategies. Natural compounds derived from plants have emerged as potential strategies for preventing cancer and treatment because of their inherent pharmacological properties. This comprehensive review aimed to evaluate the therapeutic potential of five key natural compounds: apigenin, quercetin, piperine, curcumin, and resveratrol in cancer prevention and therapy. By examining their molecular mechanisms and preclinical evidence, this review seeks to elucidate their role as potential adjuvants or stand-alone therapies in cancer management. The exploration of natural compounds as cancer therapeutics offers several advantages, including low toxicity, wide availability, and compatibility with conventional chemotherapeutic agents. We highlighted the current understanding of their anticancer mechanisms and clinical applications for advancing personalized cancer care to improve patient outcomes. We discussed the empirical findings from in vitro, in vivo, and clinical studies reporting biological activity and therapeutic efficacy in antioxidant, immunomodulatory, anti-carcinogenic, and chemo-sensitizing modes. Innovative delivery systems and personalized treatment approaches may further enhance their bioavailability and therapeutic utility in a synergistic approach with chemo- and radiotherapeutic disease management. This review underscores the importance of natural compounds in cancer prevention and treatment, promoting a multidisciplinary approach to the development of innovative therapeutic strategies.

| [1] |
Pan SY, Litscher G, Gao SH, et al. (2014) Historical perspective of traditional indigenous medical practices: The current renaissance and conservation of herbal resources. Evid Based Complement Alternat Med 2014: e525340. https://doi.org/10.1155/2014/525340 
|
| [2] |
Atanasov AG, Zotchev SB, Dirsch VM, et al. (2021) Natural products in drug discovery: Advances and opportunities. Nat Rev Drug Discov 20: 200-216. https://doi.org/10.1038/s41573-020-00114-z
|
| [3] |
Di Napoli R, Balzano N, Mascolo A, et al. (2023) What is the role of nutraceutical products in cancer patients? A systematic review of randomized clinical trials. Nutrients 15: 3249. https://doi.org/10.3390/nu15143249
|
| [4] |
Chemjong M, Kumari Yadav NA, Sarkate A, et al. (2021) Bioactive compounds, types, stability and health benefits. Plant Archives 21: 1863-1869. https://doi.org/10.51470/PLANTARCHIVES.2021.v21.S1.300
|
| [5] |
Asma ST, Acaroz U, Imre K, et al. (2022) Natural products/bioactive compounds as a source of anticancer drugs. Cancers 14: 6203. https://doi.org/10.3390/cancers14246203
|
| [6] |
Sharma D, Kapri A, Kumar S, et al. (2022) Review on Anti-tumor activity, Immunomodulatory and neuromodulatory properties of Piper longum Linn. J Ayu Herb Med 8: 55-60. https://doi.org/10.31254/jahm.2022.8112
|
| [7] |
Wu J, Li Y, He Q, et al. (2023) Exploration of the use of natural compounds in combination with chemotherapy drugs for tumor treatment. Molecules 28: 1022. https://doi.org/10.3390/molecules28031022
|
| [8] |
Auti A, Alessio N, Ballini A, et al. (2022) Protective effect of resveratrol against hypoxia-induced neural oxidative stress. JPM 12: 1202. https://doi.org/10.3390/jpm12081202
|
| [9] |
Boccellino M, Donniacuo M, Bruno F, et al. (2019) Protective effect of piceatannol and bioactive stilbene derivatives against hypoxia-induced toxicity in H9c2 cardiomyocytes and structural elucidation as 5-LOX inhibitors. Eur J Med Chem 180: 637-647. https://doi.org/10.1016/j.ejmech.2019.07.033
|
| [10] |
Coussens LM, Werb Z (2002) Inflammation and cancer. Nature 420: 860-867. https://doi.org/10.1038/nature01322
|
| [11] |
Boccellino M, D'Angelo S (2020) Anti-obesity effects of polyphenol intake: Current status and future possibilities. Int J Mol Sci 21: 5642. https://doi.org/10.3390/ijms21165642
|
| [12] |
Hou J, Karin M, Sun B (2021) Targeting cancer-promoting inflammation—Have anti-inflammatory therapies come of age?. Nat Rev Clin Oncol 18: 261-279. https://doi.org/10.1038/s41571-020-00459-9
|
| [13] |
Leslie K, Lang C, Devgan G, et al. (2006) Cyclin D1 is transcriptionally regulated by and required for transformation by activated signal transducer and activator of transcription. Cancer Res 66: 2544-2552. https://doi.org/10.1158/0008-5472.CAN-05-2203
|
| [14] |
Ji Z, He L, Regev A, et al. (2019) Inflammatory regulatory network mediated by the joint action of NF-kB, STAT3, and AP-1 factors is involved in many human cancers. Proc Natl Acad Sci USA 116: 9453-9462. https://doi.org/10.1073/pnas.1821068116
|
| [15] |
Turkson J, Bowman T, Garcia R, et al. (1998) Stat3 activation by Src induces specific gene regulation and is required for cell transformation. Mol Cell Biol 18: 2545-2552. https://doi.org/10.1128/MCB.18.5.2545
|
| [16] |
Iliopoulos D, Hirsch HA, Struhl K (2009) An epigenetic switch involving NF-κB, Lin28, Let-7 MicroRNA, and IL6 links inflammation to cell transformation. Cell 139: 693-706. https://doi.org/10.1016/j.cell.2009.10.014
|
| [17] |
Jia D, Li L, Andrew S, et al. (2017) An autocrine inflammatory forward-feedback loop after chemotherapy withdrawal facilitates the repopulation of drug-resistant breast cancer cells. Cell Death Dis 8: e2932. https://doi.org/10.1038/cddis.2017.319
|
| [18] |
Lázár-Molnár E, Hegyesi H, Tóth S, et al. (2000) Autocrine and paracrine regulation by cytokines and growth factors in melanoma. Cytokine 12: 547-554. https://doi.org/10.1006/cyto.1999.0614
|
| [19] |
Lebman DA, Edmiston JS (1999) The role of TGF-Beta in growth, differentiation, and maturation of B lymphocytes. Microbes Infect 1: 1297-1304. https://doi.org/10.1016/s1286-4579(99)00254-3
|
| [20] |
Li MO, Wan YY, Sanjabi S, et al. (2006) Transforming growth factor-beta regulation of immune responses. Annu Rev Immunol 24: 99-146. https://doi.org/10.1146/annurev.immunol.24.021605.090737
|
| [21] |
Xu J, Lamouille S, Derynck R (2009) TGF-beta-induced epithelial to mesenchymal transition. Cell Res 19: 156-172. https://doi.org/10.1038/cr.2009.5
|
| [22] |
Aggarwal BB, Takada Y, Oommen OV (2004) From chemoprevention to chemotherapy: Common targets and common goals. Expert Opin Investig Drugs 13: 1327-1338. https://doi.org/10.1517/13543784.13.10.1327
|
| [23] |
Maresso KC, Tsai KY, Brown PH, et al. (2015) Molecular cancer prevention: Current status & future directions. CA Cancer J Clin 65: 345-383. https://doi.org/10.3322/caac.21287
|
| [24] |
Gravitz L (2011) Chemoprevention: First line of defence. Nature 471: S5-S7. https://doi.org/10.1038/471S5a
|
| [25] |
G MS, Swetha M, Keerthana CK, et al. (2022) Cancer chemoprevention: A strategic approach using phytochemicals. Front Pharmacol 12: 809308. https://doi.org/10.3389/fphar.2021.809308
|
| [26] |
Fisher B, Costantino JP, Wickerham DL, et al. (1998) Tamoxifen for prevention of breast cancer: Report of the national surgical adjuvant breast and bowel project P-1 study. J Natl Cancer Inst 90: 1371-1388. https://doi.org/10.1093/jnci/90.18.1371
|
| [27] |
Vogel VG, Costantino JP, Wickerham DL, et al. (2006) Effects of tamoxifen vs raloxifene on the risk of developing invasive breast cancer and other disease outcomes: The NSABP study of tamoxifen and raloxifene (STAR) P-2 trial. JAMA 295: 2727-2741. https://doi.org/10.1001/jama.295.23.joc60074
|
| [28] |
Pruthi S, Heisey RE, Bevers TB (2015) Chemoprevention for breast cancer. Ann Surg Oncol 22: 3230-3235. https://doi.org/10.1245/s10434-015-4715-9
|
| [29] |
Higurashi T, Hosono K, Takahashi H, et al. (2016) Metformin for chemoprevention of metachronous colorectal adenoma or polyps in post-polypectomy patients without diabetes: A multicentre double-blind, placebo-controlled, randomised phase 3 trial. The Lancet Oncology 17: 475-483. https://doi.org/10.1016/S1470-2045(15)00565-3
|
| [30] |
Janicko M, Drazilova S, Pella D, et al. (2016) Pleiotropic effects of statins in the diseases of the liver. World J Gastroenterol 22: 6201-6213. https://doi.org/10.3748/wjg.v22.i27.6201
|
| [31] |
Kim G, Kang ES (2019) Prevention of hepatocellular carcinoma by statins: Clinical evidence and plausible mechanisms. Semin Liver Dis 39: 141-152. https://doi.org/10.1055/s-0039-1679956
|
| [32] |
Wang ST, Ho HJ, Lin JT, et al. (2017) Simvastatin-induced cell cycle arrest through inhibition of STAT3/SKP2 axis and activation of AMPK to promote P27 and P21 accumulation in hepatocellular carcinoma cells. Cell Death Dis 8: e2626. https://doi.org/10.1038/cddis.2016.472
|
| [33] |
Cuzick J, Otto F, Baron JA, et al. (2009) Aspirin and non-steroidal anti-inflammatory drugs for cancer prevention: An international consensus statement. Lancet Oncol 10: 501-507. https://doi.org/10.1016/S1470-2045(09)70035-X
|
| [34] |
Wong CC, Baum J, Silvestro A, et al. (2020) Inhibition of IL1β by canakinumab may be effective against diverse molecular subtypes of lung cancer: An exploratory analysis of the CANTOS trial. Cancer Res 80: 5597-5605. https://doi.org/10.1158/0008-5472.CAN-19-3176
|
| [35] |
Garlanda C, Mantovani A (2021) Interleukin-1 in tumor progression, therapy, and prevention. Cancer Cell 39: 1023-1027. https://doi.org/10.1016/j.ccell.2021.04.011
|
| [36] |
Tsai HJ (2015) Clinical cancer chemoprevention: From the Hepatitis B Virus (HBV) vaccine to the Human Papillomavirus (HPV) vaccine. Taiwan J Obstet Gynecol 54: 112-115. https://doi.org/10.1016/j.tjog.2013.11.009
|
| [37] |
Enokida T, Moreira A, Bhardwaj N (2021) Vaccines for immunoprevention of cancer. J Clin Invest 131: e146956. https://doi.org/10.1172/JCI146956
|
| [38] |
Hu Y, Yu H (2020) Prevention strategies of mother-to-child transmission of Hepatitis B Virus (HBV) infection. Pediatr Investig 4: 133-137. https://doi.org/10.1002/ped4.12205
|
| [39] |
D'Angelo S, La Porta R, Napolitano M, et al. (2012) Effect of Annurca apple polyphenols on human HaCaT keratinocytes proliferation. J Med Food 15: 1024-1031. https://doi.org/10.1089/jmf.2012.0076
|
| [40] |
Mazurakova A, Koklesova L, Samec M, et al. (2022) Anti-breast cancer effects of phytochemicals: Primary, secondary, and tertiary care. EPMA Journal 13: 315-334. https://doi.org/10.1007/s13167-022-00277-2
|
| [41] |
Mitsiogianni M, Koutsidis G, Mavroudis N, et al. (2019) The role of isothiocyanates as cancer chemo-preventive, chemo-therapeutic and anti-melanoma agents. Antioxidants (Basel) 8: 106. https://doi.org/10.3390/antiox8040106
|
| [42] |
Arrigoni R, Ballini A, Santacroce L, et al. (2022) Another look at dietary polyphenols: Challenges in cancer prevention and treatment. Curr Med Chem 29: 1061-1082. https://doi.org/10.2174/0929867328666210810154732
|
| [43] | Di Domenico M, Feola A, Ambrosio P, et al. (2020) Antioxidant effect of beer polyphenols and their bioavailability in dental-derived stem cells (D-dSCs) and human intestinal epithelial lines (Caco-2) cells. Stem Cells Int 10: 8835813. https://doi.org/10.1155/2020/8835813 |
| [44] |
Ricci S, Pinto F, Auletta A, et al. (2019) The enigmatic role of matrix metalloproteinases in epithelial-to-mesenchymal transition of oral squamous cell carcinoma: Implications and nutraceutical aspects. J Cell Biochem 120: 6813-6819. https://doi.org/10.1002/jcb.26905
|
| [45] |
Subramaniam S, Selvaduray KR, Radhakrishnan AK (2019) Bioactive compounds: Natural defense against cancer?. Biomolecules 9: 758. https://doi.org/10.3390/biom9120758
|
| [46] |
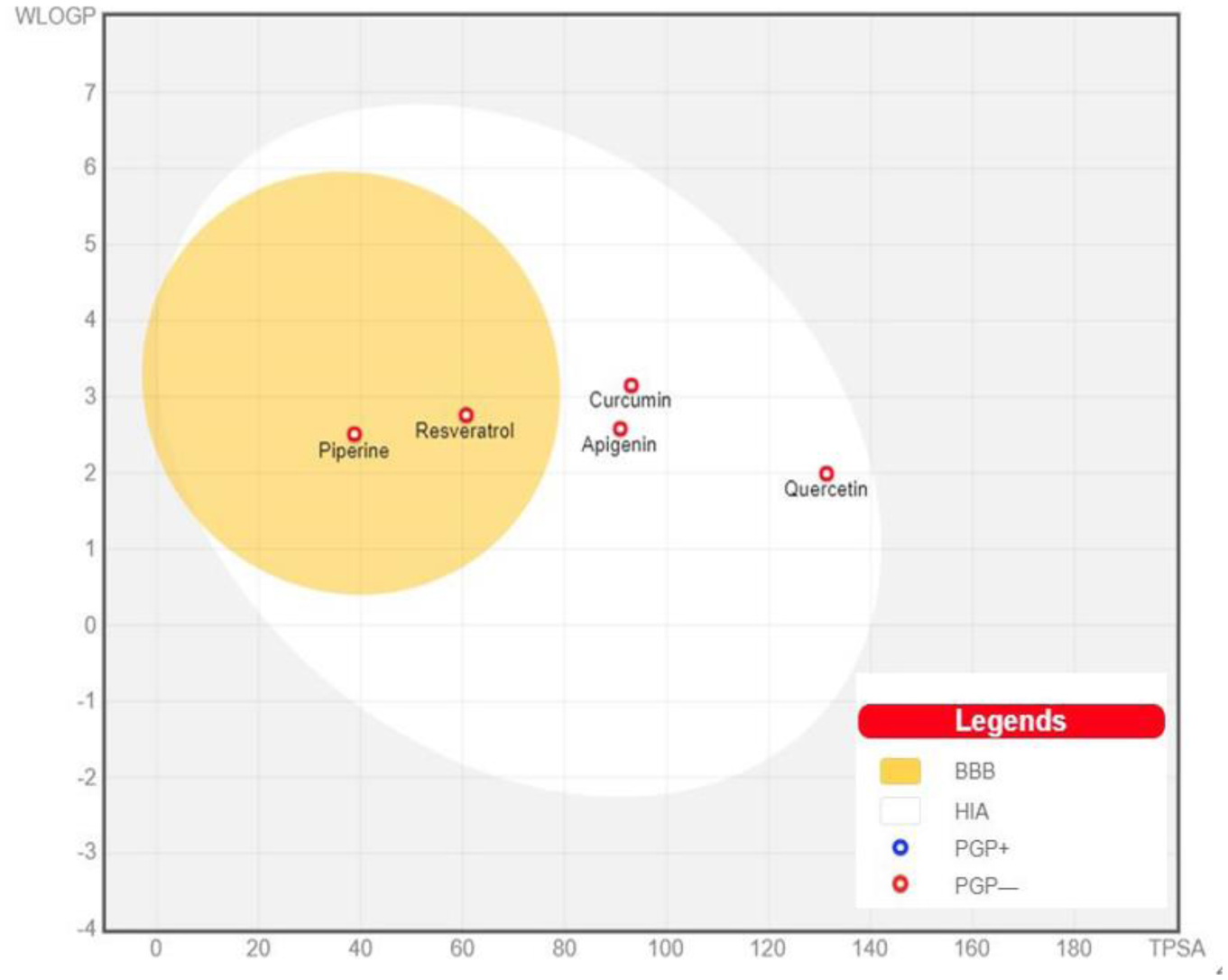
Daina A, Zoete V (2016) A BOILED-egg to predict gastrointestinal absorption and brain penetration of small molecules. Chem Med Chem 11: 1117-1121. https://doi.org/10.1002/cmdc.201600182
|
| [47] |
Daina A, Michielin O, Zoete V (2017) SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci Rep 7: 42717. https://doi.org/10.1038/srep42717
|
| [48] |
Abbott NJ, Patabendige AAK, Dolman DEM, et al. (2010) Structure and function of the blood–brain barrier. Neurobiol Dis 37: 13-25. https://doi.org/10.1016/j.nbd.2009.07.030
|
| [49] |
Sahin K, Pala R, Tuzcu M, et al. (2016) Curcumin prevents muscle damage by regulating NF-κB and Nrf2 pathways and improves performance: An in vivo model. J Inflamm Res 9: 147-154. https://doi.org/10.2147/JIR.S110873
|
| [50] |
Saberi-Karimian M, Katsiki N, Caraglia M, et al. (2019) Vascular endothelial growth factor: An important molecular target of curcumin. Crit Rev Food Sci Nutr 59: 299-312. https://doi.org/10.1080/10408398.2017.1366892
|
| [51] |
Fattori V, Pinho-Ribeiro FA, Borghi SM, et al. (2015) Curcumin inhibits superoxide anion-induced pain-like behavior and leukocyte recruitment by increasing Nrf2 expression and reducing NF-κB Activation. Inflamm Res 64: 993-1003. https://doi.org/10.1007/s00011-015-0885-y
|
| [52] |
Dai W, Wang H, Fang J, et al. (2018) Curcumin provides neuroprotection in model of traumatic brain injury via the Nrf2-ARE signaling pathway. Brain Res Bull 140: 65-71. https://doi.org/10.1016/j.brainresbull.2018.03.020
|
| [53] |
Huang T, Zhao J, Guo D, et al. (2018) Curcumin mitigates axonal injury and neuronal cell apoptosis through the PERK/Nrf2 signaling pathway following diffuse axonal injury. Neuroreport 29: 661-677. https://doi.org/10.1097/WNR.0000000000001015
|
| [54] | Zhang C, He LJ, Ye HZ, et al. (2018) Nrf2 is a key factor in the reversal effect of curcumin on multidrug resistance in the HCT-8/5-Fu human colorectal cancer cell line. Mol Med Rep 18: 5409-5416. https://doi.org/10.3892/mmr.2018.9589 |
| [55] |
Crocetto F, Boccellino M, Barone B, et al. (2020) The crosstalk between prostate cancer and microbiota inflammation: Nutraceutical products are useful to balance this interplay?. Nutrients 12: 2648. https://doi.org/10.3390/nu12092648
|
| [56] |
Lu KH, Lu PWA, Lu EWH, et al. (2023) Curcumin and its analogs and carriers: Potential therapeutic strategies for human osteosarcoma. Int J Biol Sci 19: 1241-1265. https://doi.org/10.7150/ijbs.80590
|
| [57] |
Luthra PM, Kumar R, Prakash A (2009) Demethoxycurcumin induces Bcl-2 mediated G2/M arrest and apoptosis in human glioma U87 cells. Biochem Biophys Res Commun 384: 420-425. https://doi.org/10.1016/j.bbrc.2009.04.149
|
| [58] |
Marcu MG, Jung YJ, Lee S, et al. (2006) Curcumin is an inhibitor of P300 histone acetylatransferase. Med Chem 2: 169-174. https://doi.org/10.2174/157340606776056133
|
| [59] |
Vajragupta O, Boonchoong P, Morris GM, et al. (2005) Active site binding modes of curcumin in HIV-1 protease and integrase. Bioorg Med Chem Lett 15: 3364-3368. https://doi.org/10.1016/j.bmcl.2005.05.032
|
| [60] |
Hewlings SJ, Kalman DS (2017) Curcumin: A review of its' effects on human health. Foods 6: 92. https://doi.org/10.3390/foods6100092
|
| [61] |
Shoba G, Joy D, Joseph T, et al. (1998) Influence of piperine on the pharmacokinetics of curcumin in animals and human volunteers. Planta Med 64: 353-356. https://doi.org/10.1055/s-2006-957450
|
| [62] |
Banji D, Banji OJF, Srinivas K (2021) Neuroprotective effect of turmeric extract in combination with its essential oil and enhanced brain bioavailability in an animal model. Biomed Res Int 2021: 6645720. https://doi.org/10.1155/2021/6645720
|
| [63] |
Donadio V, Incensi A, Rizzo G, et al. (2022) The effect of curcumin on idiopathic parkinson disease: A clinical and skin biopsy study. J Neuropathol Exp Neurol 81: 545-552. https://doi.org/10.1093/jnen/nlac034
|
| [64] |
Huppke P, Weissbach S, Church JA, et al. (2017) Activating de novo mutations in NFE2L2 encoding NRF2 cause a multisystem disorder. Nat Commun 8: 818. https://doi.org/10.1038/s41467-017-00932-7
|
| [65] |
Polimeno L, Viggiani MT, Giorgio F, et al. (2022) Possible role of nuclear factor erythroid 2-related factor 2 in the progression of human colon precancerous lesions. Dig Liver Dis 54: 1716-1720. https://doi.org/10.1016/j.dld.2022.01.131
|
| [66] |
Shahcheraghi SH, Salemi F, Peirovi N, et al. (2021) Nrf2 regulation by curcumin: Molecular aspects for therapeutic prospects. Molecules 27: 167. https://doi.org/10.3390/molecules27010167
|
| [67] |
Hwang D B, Won D H, Shin Y S, et al. (2020) Ccrn4l as a pre-dose marker for prediction of cisplatin-induced hepatotoxicity susceptibility. Free Radic Biol Med 148: 128-139. https://doi.org/10.1016/j.freeradbiomed.2020.01.003
|
| [68] |
Boccellino M, Ambrosio P, Ballini A, et al. (2022) The role of curcumin in prostate cancer cells and derived spheroids. Cancers (Basel) 14: 3348. https://doi.org/10.3390/cancers14143348
|
| [69] | Barinda AJ, Arozal W, Sandhiutami NMD, et al. (2022) Curcumin prevents epithelial-to mesenchymal transition-mediated ovarian cancer progression through NRF2/ETBR/ET-1 axis and preserves mitochondria biogenesis in kidney after cisplatin administration. Adv Pharm Bull 12: 128-141. https://doi.org/10.34172/apb.2022.014 |
| [70] |
Thongchot S, Jirapongwattana N, Luangwattananun P, et al. (2022) Adoptive transfer of anti-nucleolin T cells combined with PD-L1 inhibition against triple-negative breast cancer. Mol Cancer Ther 21: 727-739. https://doi.org/10.1158/1535-7163.MCT-21-0823
|
| [71] |
Ham IH, Wang L, Lee D, et al. (2022) Curcumin inhibits the cancer‑associated fibroblast‑derived chemoresistance of gastric cancer through the suppression of the JAK/STAT3 signaling pathway. Int J Oncol 61: 85. https://doi.org/10.3892/ijo.2022.5375
|
| [72] |
Ryan JL, Heckler CE, Ling M, et al. (2013) Curcumin for radiation dermatitis: A randomized, double-blind, placebo-controlled clinical trial of thirty breast cancer patients. Radiat Res 180: 34-43. https://doi.org/10.1667/RR3255.1
|
| [73] |
Sahebkar A, Cicero AFG, Simental-Mendía LE, et al. (2016) Curcumin downregulates human tumor necrosis factor-α levels: A systematic review and meta-analysis ofrandomized controlled trials. Pharmacol Res 107: 234-242. https://doi.org/10.1016/j.phrs.2016.03.026
|
| [74] |
Hidayat YM, Wagey F, Suardi D, et al. (2021) Analysis of curcumin as a radiosensitizer in cancer therapy with serum survivin examination: Randomised control trial. Asian Pac J Cancer Prev 22: 139-143. https://doi.org/10.31557/APJCP.2021.22.1.139
|
| [75] |
Ashrafizadeh M, Bakhoda MR, Bahmanpour Z, et al. (2020) Apigenin as tumor suppressor in cancers: Biotherapeutic activity, nanodelivery, and mechanisms with emphasis on pancreatic cancer. Front Chem 8: 829. https://doi.org/10.3389/fchem.2020.00829
|
| [76] |
Yan X, Qi M, Li P, et al. (2017) Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci 7: 50. https://doi.org/10.1186/s13578-017-0179-x
|
| [77] |
Javed Z, Sadia H, Iqbal MJ, et al. (2021) Apigenin role as cell-signaling pathways modulator: Implications in cancer prevention and treatment. Cancer Cell Int 21: 189. https://doi.org/10.1186/s12935-021-01888-x
|
| [78] |
Zhang C, Liao Y, Li T, et al. (2024) Apigenin promotes apoptosis of 4T1 cells through PI3K/AKT/Nrf2 pathway and improves tumor immune microenvironment in vivo. Toxicol Res (Camb) 13: tfae011. https://doi.org/10.1093/toxres/tfae011
|
| [79] |
Sudhakaran M, Navarrete TG, Mejía-Guerra K, et al. (2023) Transcriptome reprogramming through alternative splicing triggered by apigenin drives cell death in triple-negative breast cancer. Cell Death Dis 14: 824. https://doi.org/10.1038/s41419-023-06342-6
|
| [80] |
Bonilla-Vidal L, Świtalska M, Espina M, et al. (2023) Dually active apigenin-loaded nanostructured lipid carriers for cancer treatment. Int J Nanomedicine 18: 6979-6997. https://doi.org/10.2147/IJN.S429565
|
| [81] |
Yang B, Mao Y, Zhang Y, et al. (2023) HA-coated PLGA nanoparticles loaded with apigenin for colon cancer with high expression of CD44. Molecules 28: 7565. https://doi.org/10.3390/molecules28227565
|
| [82] |
Zhu ZY, Gao T, Huang Y, et al. (2016) Apigenin ameliorates hypertension-induced cardiac hypertrophy and down-regulates cardiac hypoxia inducible factor-Lα in rats. Food Funct 7: 1992-1998. https://doi.org/10.1039/c5fo01464f
|
| [83] | Giudice A, Montella M, Boccellino M, et al. (2017) Epigenetic changes induced by green tea catechins are associated with prostate cancer. Curr Mol Med 17: 405-420. https://doi.org/10.2174/1566524018666171219101937 |
| [84] |
Mirzoeva S, Franzen CA, Pelling JC (2014) Apigenin inhibits TGF-β-induced VEGF expression in human prostate carcinoma cells via a smad2/3- and Src-dependent mechanism. Mol Carcinog 53: 598-609. https://doi.org/10.1002/mc.22005
|
| [85] |
Melstrom LG, Salabat MR, Ding XZ, et al. (2011) Apigenin down-regulates the hypoxia response genes: HIF-1α, GLUT-1, and VEGF in human pancreatic cancer cells. J Surg Res 167: 173-181. https://doi.org/10.1016/j.jss.2010.10.041
|
| [86] |
Turkekul K, Erdogan S (2023) Potent suppression of prostate cancer cell growth and eradication of cancer stem cells by CD44-targeted nanoliposome-quercetin nanoparticles. J Cancer Prev 28: 160-174. https://doi.org/10.15430/JCP.2023.28.4.160
|
| [87] |
Isacco CG, Ballini A, De Vito D, et al. (2021) Rebalancing the oral microbiota as an efficient tool in endocrine, metabolic and immune disorders. Endocr Metab Immune Disord Drug Targets 21: 777-784. https://doi.org/10.2174/1871530320666200729142504
|
| [88] | Ballini A, Scacco S, Boccellino M, et al. (2020) Microbiota and obesity: Where are we now?. Biology (Basel) 9: 415. https://doi.org/10.3390/biology9120415 |
| [89] |
Di Domenico M, Ballini A, Boccellino M, et al. (2022) The intestinal microbiota may be a potential theranostic tool for personalized medicine. J Pers Med 12: 523. https://doi.org/10.3390/jpm12040523
|
| [90] |
Marino MM, Nastri BM, D'Agostino M, et al. (2022) Does gut-breast microbiota axis orchestrates cancer progression?. Endocr Metab Immune Disord Drug Targets 22: 1111-1122. https://doi.org/10.2174/1871530322666220331145816
|
| [91] |
Wu R, Xiong J, Zhou T, et al. (2023) Quercetin/anti-PD-1 antibody combination therapy regulates the gut microbiota, impacts macrophage immunity and reshapes the hepatocellular carcinoma tumor microenvironment. Front Biosci (Landmark Ed) 28: 327. https://doi.org/10.31083/j.fbl2812327
|
| [92] |
Topçu-Tarladaçalışır Y, Sapmaz-Metin M, Mercan Z, et al. (2024) Quercetin attenuates endoplasmic reticulum stress and apoptosis in TNBS-induced colitis by inhibiting the glucose regulatory protein 78 activation. Balkan Med J 41: 30-37. https://doi.org/10.4274/balkanmedj.galenos.2023.2023-10-9
|
| [93] |
DiGuilio KM, Rybakovsky E, Valenzano MC, et al. (2023) Quercetin improves and protects calu-3 airway epithelial barrier function. Front Cell Dev Biol 11: 1271201. https://doi.org/10.3389/fcell.2023.1271201
|
| [94] |
Asl AM, Kalaee M, Abdouss M, et al. (2024) Novel targeted delivery of quercetin for human hepatocellular carcinoma using starch/polyvinyl alcohol nanocarriers based hydrogel containing Fe2O3 nanoparticles. Int J Biol Macromol 257: 128626. https://doi.org/10.1016/j.ijbiomac.2023.128626
|
| [95] |
Xu YH, Xu JB, Chen LL, et al. (2023) Protective mechanisms of quercetin in neonatal rat brain injury induced by Hypoxic-Ischemic Brain Damage (HIBD). Food Sci Nutr 11: 7649-7663. https://doi.org/10.1002/fsn3.3684
|
| [96] | Zhang N, Qiu Q, Chen Y, et al. (2023) Quercetin improves pulmonary arterial hypertension in rats by regulating the HMGB1/RAGE/NF-κB pathway. Nan Fang Yi Ke Da Xue Xue Bao 43: 1606-1612. https://doi.org/10.12122/j.issn.1673-4254.2023.09.19 |
| [97] |
Rathi V, Tiwari I, Kulshreshtha R, et al. (2023) Hypobaric hypoxia induced renal injury in rats: prophylactic amelioration by quercetin supplementation. PLoS One 18: e0279304. https://doi.org/10.1371/journal.pone.0279304
|
| [98] |
Johri RK, Zutshi U (1992) An ayurvedic formulation ‘trikatu’ and its constituents. J Ethnopharmacol 37: 85-91. https://doi.org/10.1016/0378-8741(92)90067-2
|
| [99] |
Benayad S, Wahnou H, El Kebbaj R, et al. (2023) The promise of piperine in cancer chemoprevention. Cancers 15: 5488. https://doi.org/10.3390/cancers15225488
|
| [100] |
Greenshields AL, Doucette CD, Sutton KM, et al. (2015) Piperine inhibits the growth and motility of triple-negative breast cancer cells. Cancer Lett 357: 129-140. https://doi.org/10.1016/j.canlet.2014.11.017
|
| [101] |
Sun X, Xu Q, Zeng L, et al. (2020) Resveratrol suppresses the growth and metastatic potential of cervical cancer by inhibiting STAT3Tyr705 phosphorylation. Cancer Med 9: 8685-8700. https://doi.org/10.1002/cam4.3510
|
| [102] |
Rao YL, Ganaraja B, Joy T, et al. (2020) Neuroprotective effects of resveratrol in Alzheimer's disease. Front Biosci (Elite Ed) 12: 139-149. https://doi.org/10.2741/E863
|
| [103] |
Cao D, Wang M, Qiu X, et al. (2015) Structural basis for allosteric, substrate-dependent stimulation of SIRT1 activity by resveratrol. Genes Dev 29: 1316-1325. https://doi.org/10.1101/gad.265462.115
|
| [104] |
Wiciński M, Erdmann J, Nowacka A, et al. (2023) Natural phytochemicals as SIRT activators-focus on potential biochemical mechanisms. Nutrients 15: 3578. https://doi.org/10.3390/nu15163578. PMID: 37630770; PMCID: PMC10459499
|
| [105] |
Yu HP, Hwang TL, Hsieh PW, et al. (2011) Role of estrogen receptor-dependent upregulation of P38 MAPK/Heme oxygenase 1 in resveratrol-mediated attenuation of intestinal injury after trauma-hemorrhage. Shock 35: 517. https://doi.org/10.1097/SHK.0b013e318209e931
|
| [106] |
Fernández-Rodríguez JA, Almonte-Becerril M, Ramil-Gómez O, et al. (2021) Autophagy activation by resveratrol reduces severity of experimental rheumatoid arthritis. Mol Nutr Food Re 65: 2000377. https://doi.org/10.1002/mnfr.202000377
|
| [107] |
Pahl HL (1999) Activators and target genes of Rel/NF-κB transcription factors. Oncogene 18: 6853-6866. https://doi.org/10.1038/sj.onc.1203239
|
| [108] |
Ji Yu E, Kim SH, Heo K, et al. (2011) Reciprocal roles of DBC1 and SIRT1 in regulating estrogen receptor α activity and co-activator synergy. Nucleic Acids Res 39: 6932-6943. https://doi.org/10.1093/nar/gkr347
|
| [109] |
Nwachukwu JC, Srinivasan S, Bruno NE, et al. (2014) Resveratrol modulates the inflammatory response via an estrogen receptor-signal integration network. eLife 3: e02057. https://doi.org/10.7554/eLife.02057
|
| [110] |
Buhrmann C, Yazdi M, Popper B, et al. (2018) Resveratrol chemosensitizes TNF-β-induced survival of 5-FU-treated colorectal cancer cells. Nutrients 10: 888. https://doi.org/10.3390/nu10070888
|
| [111] |
Wang Y, Chen X, Tang N, et al. (2024) Boosting clear cell renal carcinoma-specific drug discovery using a deep learning algorithm and single-cell analysis. Int J Mol Sci 25: 4134. https://doi.org/10.3390/ijms25074134
|


Figures(3) / Tables(1)

Amogh Auti, Madhura Tathode, Maria Michela Marino, Antonio Vitiello, Andrea Ballini, Francesco Miele, Valeria Mazzone, Alessia Ambrosino, Mariarosaria Boccellino. Nature's weapons: Bioactive compounds as anti-cancer agents[J]. AIMS Public Health, 2024, 11(3): 747-772. doi: 10.3934/publichealth.2024038







 DownLoad:
DownLoad: