In Malaysia, breast cancer accounted for 34.1% of all female cancer cases with women presenting breast cancer at late stages. Breast cancer has a higher five-year survival rate if detected early. An increase of approximately 30% in the five-year survival rate is indicated if breast cancer is detected at stage III compared to stage IV. Thus, survival rate of breast cancer can be increased by creating awareness and encouraging breast cancer screening amongst women. Breast self-examination (BSE) is highly recommended for breast cancer screening due to its simplicity with no incurred cost. The Health Belief Model is used in this study to explain and predict the adoptive behavior of BSE amongst women in Kuantan, Pahang.
This study employed a multi-stage sampling method using a simple proportion formula at 5% type 1 error, p < 0.05 and absolute error at 2% which resulted in a sample of 520 participants. The data for the study was obtained using a validated bilingual self-constructed questionnaire and the model constructed using Mplus software.
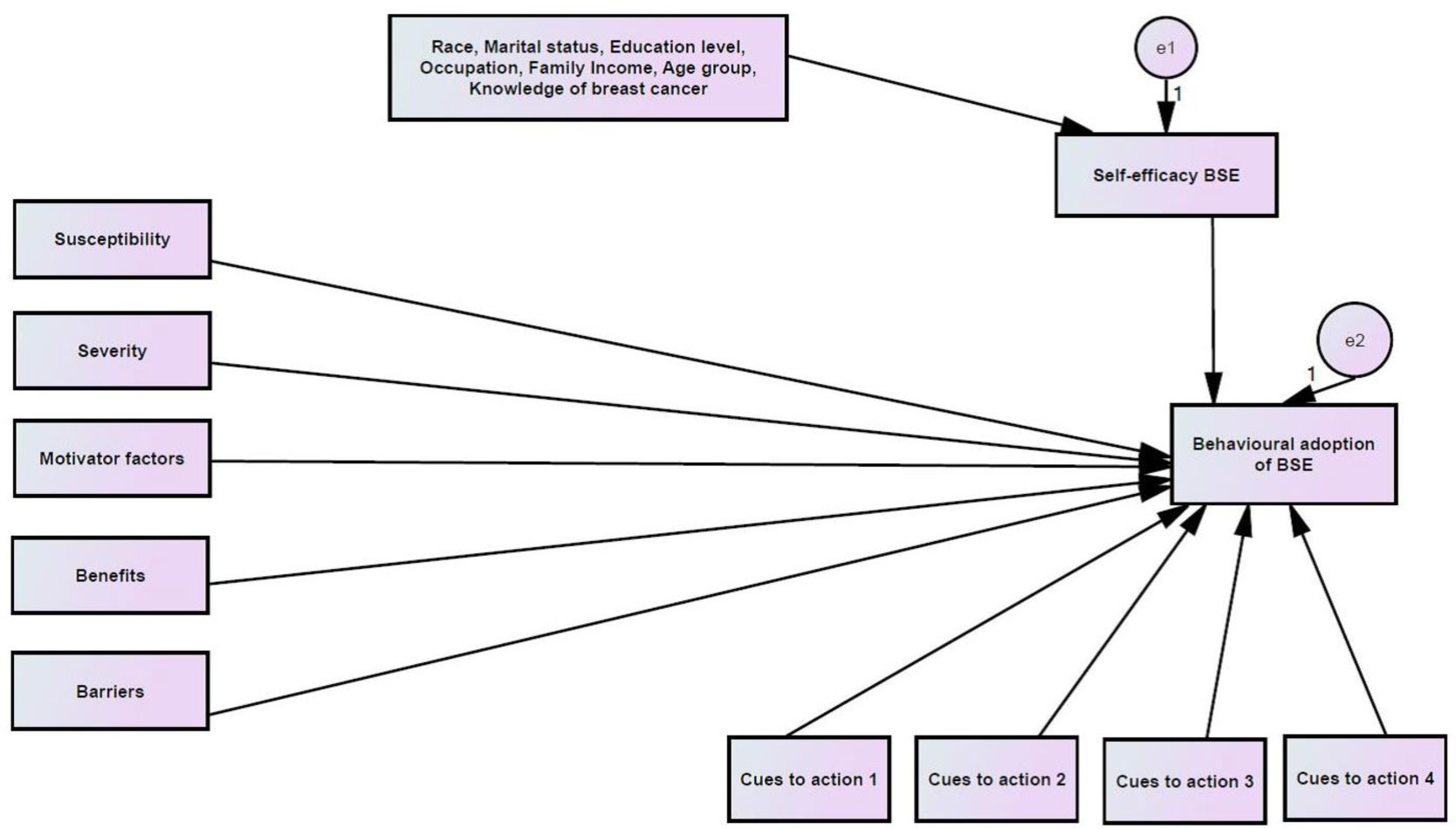
Perceived severity, benefits and barriers were found to significantly influence the behavioral adoption of BSE. Married women aged from 45 to 55 years and knowledge were found to significantly moderate the relationship between perceived benefits and behavioral adoption of BSE. Further, self-efficacy was found as the core construct that mediates the relationship between married women aged 45 to 55 years and the behavioral adoption of BSE.
Self-efficacy is found in the study to influence the behavioral adoption of BSE. This is undeniable as self-efficacy can promote confidence in initiating and maintenance of behavioral change if the perceived change is beneficial at an acceptable cost.
Citation: Soo-Foon Moey, Norfariha Che Mohamed, Bee-Chiu Lim. A path analytic model of health beliefs on the behavioral adoption of breast self-examination[J]. AIMS Public Health, 2021, 8(1): 15-31. doi: 10.3934/publichealth.2021002
In Malaysia, breast cancer accounted for 34.1% of all female cancer cases with women presenting breast cancer at late stages. Breast cancer has a higher five-year survival rate if detected early. An increase of approximately 30% in the five-year survival rate is indicated if breast cancer is detected at stage III compared to stage IV. Thus, survival rate of breast cancer can be increased by creating awareness and encouraging breast cancer screening amongst women. Breast self-examination (BSE) is highly recommended for breast cancer screening due to its simplicity with no incurred cost. The Health Belief Model is used in this study to explain and predict the adoptive behavior of BSE amongst women in Kuantan, Pahang.
This study employed a multi-stage sampling method using a simple proportion formula at 5% type 1 error, p < 0.05 and absolute error at 2% which resulted in a sample of 520 participants. The data for the study was obtained using a validated bilingual self-constructed questionnaire and the model constructed using Mplus software.
Perceived severity, benefits and barriers were found to significantly influence the behavioral adoption of BSE. Married women aged from 45 to 55 years and knowledge were found to significantly moderate the relationship between perceived benefits and behavioral adoption of BSE. Further, self-efficacy was found as the core construct that mediates the relationship between married women aged 45 to 55 years and the behavioral adoption of BSE.
Self-efficacy is found in the study to influence the behavioral adoption of BSE. This is undeniable as self-efficacy can promote confidence in initiating and maintenance of behavioral change if the perceived change is beneficial at an acceptable cost.
Breast self-examination
Health belief model
Exploratory factor analysis
Structural equation modeling

| [1] | World Cancer Research Fund International, American Institute for Cancer Research Worldwide cancer data: Global cancer statistics for the most common cancers (2020) .Available from: https://www.wcrf.org/dietandcancer/cancer-trends/worldwide-cancer-data. |
| [2] | Ministry of Health MalaysiaMalaysia National Cancer Registry Report 2012–2016. Putrajaya, Malaysia: National Cancer Institute. |
| [3] | Ferlay J, Soerjomataram I, Ervik M, et al. (2015) GLOBOCAN 2012 v1. 0, Cancer Incidence and Mortality Worldwide: IARC CancerBase No. 11. International Agency for Research on Cancer, Lyon, France. 2013. Globocan Iarc Fr . |
| [4] |
Abdullah NA, Wan Mahiyuddin W, Muhammad NA, et al. (2013) Survival rate of breast cancer patients in Malaysia: A population-based study. Asian Pac J Cancer Prev 14: 4591-4594. doi: 10.7314/APJCP.2013.14.8.4591

|
| [5] |
Ibrahim NI, Dahlui M, Aina E, et al. (2012) Who are the breast cancer survivors in Malaysia? Asian Pac J Cancer Prev 13: 2213-2218. doi: 10.7314/APJCP.2012.13.5.2213
|
| [6] |
Bhoo Pathy N, Yip CH, Taib NA, et al. (2011) Breast cancer in a multi-ethnic Asian setting: Results from the Singapore-Malaysia hospital-based breast cancer registry. Breast J 20: 75-80. doi: 10.1016/j.breast.2011.01.015
|
| [7] | Kanaga KC, Nithiya J, Noor Shatirah MFV (2011) Awareness of breast cancer and screening procedures among Malaysian women. Asian Pac J Cancer Prev 12: 1965-1967. |
| [8] | Akhtari-Zavare M, Aliyan-Fini F, Ghanbari-Baghestan A, et al. (2018) Development and validation of breast cancer knowledge and beliefs questionnaire for Malaysian student population. Pertanika J Soc Sci Hum 26: 2541-2554. |
| [9] | Yip CH, Pathy NB, Teo SH (2014) A review of breast cancer research in Malaysia. Med J Malaysia 69: 7-22. |
| [10] |
Dahlui M, Eng DHG, Taib NA, et al. (2012) Predictors of breast cancer screening uptake: a pre intervention community survey in Malaysia. Asian Pac J Cancer Prev 13: 3443-3449. doi: 10.7314/APJCP.2012.13.7.3443
|
| [11] |
Al-Naggar RA, Bobryshev YV, Al-Jashamy K (2012) Practice of breast self-examination among women in Malaysia. Asian Pac J Cancer Prev 13: 3829-3833. doi: 10.7314/APJCP.2012.13.8.3829
|
| [12] |
Akhtari-Zavare M, Juni MH, Said SM, et al. (2013) Beliefs and behavior of Malaysia undergraduate female students in a public university toward breast self-examination practice. Asian Pac J Cancer Prev 14: 57-61. doi: 10.7314/APJCP.2013.14.1.57
|
| [13] |
Ghahremani L, Mousavi Z, Kaveh MH (2016) Self-care education programs based on a trans-theoretical model in women referring to health centers: breast self-examination behavior in Iran. Asian Pac J Cancer Prev 17: 5133-5138. doi: 10.7314/APJCP.2016.17.3.1157
|
| [14] |
Mohamed NC, Moey SF, Lim BC (2019) Validity and reliability of Health Belief Model questionnaire for promoting breast self-examination and screening mammogram for early cancer detection. Asian Pac J Cancer Prev 20: 2865-2873. doi: 10.31557/APJCP.2019.20.9.2865
|
| [15] | (2002) Institute of MedicineSpeaking of health: assessing health communication strategies for diverse populations. Washington D.C.: The National Academies Press 28-75. |
| [16] |
Murphy CC, Vernon SW, Diamond PM, et al. (2014) Competitive testing of health behavior theories: how do benefits, barriers, subjective norm, and intention influence mammography behavior? Ann Behav Med 47: 120-129. doi: 10.1007/s12160-013-9528-0
|
| [17] | Department of Statistics Malaysia The Source of Malaysia's Official Statistics (2020) .Available from: https://www.dosm.gov.my/v1/index.php?r=column/ctheme&menu_id=L0pheU43NWJwRWVSZklWdzQ4TlhUUT09&bul_id=MDMxdHZjWTk1SjFzTzNkRXYzcVZjdz09. |
| [18] |
Huang HT, Kuo YM, Wang SR, et al. (2016) Structural factors affecting health examination behavioral intention. Int J Environ Res Public Health 13: 395-409. doi: 10.3390/ijerph13040395
|
| [19] | Hair JF, Black WC, Babin BJ, et al. (2009) Multivariate data analysis. Exploratory factor analysis Upper Saddle River, NJ: Pearson Prentice-Hall, 90-150. |
| [20] |
Hu LT, Bentler PM (1999) Cutoff criteria for fit Indexes in covariance structure analysis: Conventional criteria versus new alternatives. Struct Equ Modeling 6: 1-55. doi: 10.1080/10705519909540118
|
| [21] | Kline RB (2011) Principles and practice of structural equation modeling New York: The Guilford Press. |
| [22] | Schermelleh-Engel K, Moosbrugger H, Muller H (2003) Evaluating the fit of structural equation models: Tests of significance and descriptive goodness-of-fit measures. Methods Psychological Res 8: 23-74. |
| [23] | Bryne BM (2010) Structural equation modeling with AMOS basic concepts, applications, and programming New York: Taylor and Francis Group, LLC. |
| [24] |
Wang J, Wang X (2012) Structural equation modeling: applications using Mplus United Kingdom: John Wiley & Sons Ltd. doi: 10.1002/9781118356258
|
| [25] |
Whittaker TA (2012) Using the modification index and standardized expected parameter change for model modification. J Exp Educ 80: 26-44. doi: 10.1080/00220973.2010.531299
|
| [26] |
Tarka P (2018) An overview of structural equation modeling: Its beginnings, historical development, usefulness and controversies in the social sciences. Qual Quant 52: 313-354. doi: 10.1007/s11135-017-0469-8
|
| [27] |
Avci IA (2008) Factors associated with breast self-examination practices and beliefs in female workers at a Muslim community. Eur J Oncol Nurs 12: 127-133. doi: 10.1016/j.ejon.2007.11.006
|
| [28] |
Noroozi A, Jomand T, Tahmasebi R (2011) Determinants of breast self-examination performance among Iranian women: An application of the Health Belief Model. J Canc Educ 26: 365-374. doi: 10.1007/s13187-010-0158-y
|
| [29] |
Maddux JE, Rogers RW (1983) Protection motivation and self-efficacy: a revised theory of fear appeals and attitude change. J Exp Social Psychol 19: 469-479. doi: 10.1016/0022-1031(83)90023-9
|
| [30] |
Beatson R, McLennan J (2011) What applied social psychology theories might contribute to community bushfire safety research after Victoria's ‘Black Saturday’? Aust Psychol 46: 171-182. doi: 10.1111/j.1742-9544.2011.00041.x
|
| [31] |
Westcott R, Ronan K, Bambrick H, et al. (2017) Expanding protection motivation theory: investigating an application to animal owners and emergency responders in bushfire emergencies. BMC Psychol 5: 1-14. doi: 10.1186/s40359-017-0182-3
|
| [32] |
Paton D, Johnston D (2001) Disasters and communities: vulnerability, resilience and preparedness. Disaster Prev Manag 10: 270-277. doi: 10.1108/EUM0000000005930
|
| [33] |
Paton D (2013) Disaster resilient communities: developing and testing an all-hazards theory. J Integr Disaster Risk Manag 3: 1-17. doi: 10.5595/idrim.2013.0050
|
| [34] | Heidari Z, Mahmoudzadeh-Sagheb HR, Sakhavar N (2008) Breast cancer screening knowledge and practice among women in southeast of Iran. Acta Med Iran 46: 321-328. |
| [35] |
Tavafian SS, Hasani L, Aghamolaei T, et al. (2009) Prediction of breast self-examination in a sample of Iranian women: an application of the Health Belief Model. BMC Womens Health 9: 37-43. doi: 10.1186/1472-6874-9-37
|
| [36] |
Kessler TA (2012) Increasing mammography and cervical cancer knowledge and screening behaviors with an educational program. Oncol Nurs Forum 39: 61-68. doi: 10.1188/12.ONF.61-68
|
| [37] |
Lotfi B, Hashemi SZ, Ansari-Moghadam A (2011) Investigation of the impact of HBM-based training on BSE in women referred to health centers in Zahedan in 2010–2011. J Health Scope 1: 39-43. doi: 10.5812/jhs.4698
|
| [38] | Khiyali Z, Aliyan F, Kashfi SH, et al. (2017) Educational intervention on breast self-examination behavior in women referred to health centers: application of Health Belief Model. Asian Pac J Cancer Prev 18: 2833-2838. |
| [39] | Demirkiran F, Ozgun H, Eskin M, et al. (2011) Cognition of breast cancer among gestational age Turkish women: a cross-sectional study. Asian Pac J Cancer Prev 12: 277-282. |
| [40] |
Elobaid YE, Aw TC, Grivna M, et al. (2014) Breast cancer screening awareness, knowledge, and practice among Arab women in the United Arab Emirates: a cross-sectional survey. PloS One 9: e105783. doi: 10.1371/journal.pone.0105783
|
| [41] |
Moey SF, Mutalib AMA, Mohamed NC, et al. (2020) The relationship of socio-demographic characteristics and knowledge of breast cancer on stage of behavioral adoption of breast self-examination. AIMS Public Health 7: 620-633. doi: 10.3934/publichealth.2020049
|
| [42] |
Akhtari-Zavare M, Juni MH, Ismail IZ, et al. (2015) Barriers to breast self-examination practice among Malaysian female students: a cross-sectional study. Springerplus 4: 692-697. doi: 10.1186/s40064-015-1491-8
|
| [43] |
Liu YL, Wang DW, Yang ZC, et al. (2019) Marital status is an independent prognostic factor in inflammatory breast cancer patients: an analysis of the surveillance, epidemiology, and end results database. Breast Cancer Res Treat 178: 379-88. doi: 10.1007/s10549-019-05385-8
|
| [44] |
Kirag N, Klzllkaya M (2019) Application of the Champion Health Belief Model to determine beliefs and behaviors of Turkish women academicians regarding breast cancer screening: a cross sectional descriptive study. BMC Womens Health 19: 1-10. doi: 10.1186/s12905-019-0828-9
|
| [45] |
Dewi TK, Massar K, Ruiter RAC, et al. (2019) Determinants of breast self-examination practice among women in Surabaya, Indonesia: An application of the health belief model. BMC Public Health 19: 1-8. doi: 10.1186/s12889-018-6343-3
|
| [46] |
Didarloo A, Nabilou B, Khalkhali HR (2017) Psychosocial predictors of breast self-examination behavior among female students: an application of the health belief model using logistic regression. BMC Public Health 17: 1-8. doi: 10.1186/s12889-017-4880-9
|
| [47] | Glanz K, Rimer BK, Viswanath K (2008) Health behavior and health education: theory, research, and practice USA, San Francisco: Jossey-Bass Inc Pub. |
| [48] |
Alkhasawneh IM (2007) Knowledge and practice of breast cancer screening among Jordanian nurses. Oncol Nurs Forum 34: 1211-1217. doi: 10.1188/07.ONF.1211-1217
|
| [49] |
Abolfotouh MA, Banimustafa AA, Mahfouz AA, et al. (2015) Using the health belief model to predict breast self-examination among Saudi women. BMC Public Health 15: 1-12. doi: 10.1186/s12889-015-2510-y
|
| [50] | Minhat H, Mustafa J, Mohd Zain N (2014) The practice of breast self-examination (BSE) among women living in an urban setting in Malaysia. Int J Public Health Clin Sci 1: 91-99. |
| [51] | Sama CB, Dzekem B, Kehbila J, et al. (2017) Awareness of breast cancer and breast self-examination among female undergraduate students in a higher teachers training college in Cameroon. Pan Afr Med J 28: 91. |
| [52] |
Gomez SL, Quach T, Horn-Ross PL, et al. (2010) Hidden breast cancer disparities in Asian women: disaggregating incidence rates by ethnicity and migrant status. Am J Public Health 100: S125-S131. doi: 10.2105/AJPH.2009.163931
|
| [53] |
Rendall MS, Weden MM, Favreault MM, et al. (2011) The protective effect of marriage for survival: a review and update. Demography 48: 481-506. doi: 10.1007/s13524-011-0032-5
|
| [54] |
Elmore L, Deshpande A, Daly M, et al. (2015) Postmastectomy radiation therapy in T3 node-negative breast cancer. J Surg Res 199: 90-96. doi: 10.1016/j.jss.2015.04.012
|
| [55] | Dewi TK, Zein RA (2017) Predicting intention perform breast self-examination: application of the Theory of Reasoned Action. Asian Pac J Cancer Prev 18: 2945-2952. |
| [56] |
Aljohani S, Saib I, Noorelahi M (2017) Women performance of breast cancer screening (Breast Self-Examination, Clinical Breast Exam and Mammography). Adv Breast Cancer Res 6: 16-27. doi: 10.4236/abcr.2017.61002
|
| [57] |
Yoshany N, Morowatisharifabad MA, Mihanpour H, et al. (2017) The effect of husbands' education regarding menopausal health on marital satisfaction of their wives. J Menopausal Med 23: 15-24. doi: 10.6118/jmm.2017.23.1.15
|
| [58] | Bandura A (1997) Self-efficacy: The exercise of control New York: W. H. Freeman. |
| [59] |
Barbeau K, Boileau K, Sarr F, et al. (2019) Path analysis in Mplus: A tutorial using a conceptual model of psychological and behavioral antecedents of bulimic symptoms in young adults. Quant Meth Psych 15: 38-53. doi: 10.20982/tqmp.15.1.p038
|
 publichealth-08-01-002-s001.pdf publichealth-08-01-002-s001.pdf |
 |


Figures(3) / Tables(7)

Soo-Foon Moey, Norfariha Che Mohamed, Bee-Chiu Lim. A path analytic model of health beliefs on the behavioral adoption of breast self-examination[J]. AIMS Public Health, 2021, 8(1): 15-31. doi: 10.3934/publichealth.2021002







 DownLoad:
DownLoad: