Citation: Nicola Bellomo, Raluca Eftimie, Guido Forni. What is the in-host dynamics of the SARS-CoV-2 virus? A challenge within a multiscale vision of living systems[J]. Networks and Heterogeneous Media, 2024, 19(2): 655-681. doi: 10.3934/nhm.2024029

| [1] |
J. P. Agnelli, B. Buffa, D. A. Knopoff, G. Torres, A spatial kinetic model of crowd evacuation dynamics with infectious disease contagion, Bull Math Biol, 85(2023), 23. https://doi.org/10.1007/s11538-023-01127-6 doi: 10.1007/s11538-023-01127-6

|
| [2] |
M. Aguiar, G. Dosi, D. A. Knopoff, M.E. Virgillito, A multiscale network-based model of contagion dynamics: heterogeneity, spatial distancing and vaccination, Math Models Methods Appl Sci, 31 (2021), 2425–2570. https://doi.org/10.1142/S0218202521500524 doi: 10.1142/S0218202521500524
|
| [3] |
K. G. Andersen, A. Rambaut, W. Ian Lipkin, E. C. Holmes, R. F. Garry, The proximal origin of SARS-CoV-2, Nat. Med., 26 (2020), 450–452. https://doi.org/10.1038/s41591-020-0820-9 doi: 10.1038/s41591-020-0820-9
|
| [4] |
S. Asgari, L. A. Pousaz, Human genetic variants identified that affect Covid susceptibility and severity, Nature, 600 (2021), 690–691. https://doi.org/10.1038/s41586-021-04210-x doi: 10.1038/s41586-021-04210-x
|
| [5] |
A. Atifa, M. A. Khan, K. Isakakova, F. S. Al-Duais, I. Ahmad, Mathematical modelling and analysis of the SARS-CoV-2 disease with reinfection, Comput. Biol. Chem., 98 (2022), 107678. https://doi.org/10.1016/j.compbiolchem.2022.107678 doi: 10.1016/j.compbiolchem.2022.107678
|
| [6] | B. Avishai, The pandemic isn't a black swan but a portent of a more fragile global system. The New Yorker, 2020. Available from: https://www.newyorker.com/news/daily-comment/the-pandemic-isnt-a-black-swan-but-a-portent-of-a-more-fragile-global-system |
| [7] | Y. M. Bar-On, A. Flamholz, R. Phillips, R. Milo, SARS-CoV-2 (COVID-19) by the numbers, eLife, 9, e57309, (2020). https://doi.org/10.7554/eLife.57309 |
| [8] |
N. Bellomo, R. Bingham, M. Chaplain, G. Dosi, G. Forni, D. Knopoff, et al., A multi-scale model of virus pandemic: Heterogeneous interactive entities in a globally connected world, Math Models Methods Appl Sci, 30 (2020), 1591–1651. https://doi.org/10.1142/S0218202520500323 doi: 10.1142/S0218202520500323
|
| [9] |
N. Bellomo, F. Brezzi, M. Chaplain, Modelling Virus pandemics in a globally connected world, a challenge towards a mathematics for living lystems, Math Models Methods Appl Sci, 31 (2021), 2391–2397. https://doi.org/10.1142/S0218202521020024 doi: 10.1142/S0218202521020024
|
| [10] |
N. Bellomo, D. Burini, G. Dosi, L. Gibelli, D. A. Knopoff, N. Outada, et al., What is life? A perspective of the mathematical kinetic theory of active particles, Math Models Methods Appl Sci, 31 (2021), 1821–1866. https://doi.org/10.1142/S0218202521500408 doi: 10.1142/S0218202521500408
|
| [11] |
N. Bellomo, D. Burini, N. Outada, Multiscale models of Covid-19 with mutations and variants, Netw. Heterog. Media., 17 (2022), 293–310. https://doi.org/10.3934/nhm.2022008 doi: 10.3934/nhm.2022008
|
| [12] |
N. Bellomo, D. Burini, N. Outada, Pandemics of Mutating Virus and Society: A multi-scale active particles approach, Philos. Trans. Royal Soc. A, 380 (2022), 20210161. https://doi.org/10.1098/rsta.2021.0161 doi: 10.1098/rsta.2021.0161
|
| [13] |
N. Bellomo, L. Gibelli, N. Outada, On the interplay between behavioral dynamics and social interactions in human crowds, Kinet. Relat. Models, 12 (2019), 397–409. https://doi.org/10.3934/krm.2019017 doi: 10.3934/krm.2019017
|
| [14] |
G. Bertaglia, L. Pareschi, Hyperbolic compartmental models for epidemic spread on networks with uncertain data: application to the emergence of Covid-19 in Italy, Math Models Methods Appl Sci, 31 (2021), 2495–2531. https://doi.org/10.1142/S0218202521500548 doi: 10.1142/S0218202521500548
|
| [15] | G. Bertaglia, A. Bondesan, D. Burini, R. Eftimie, L. Pareschi, G. Toscani, New trends on the systems approach to modeling SARS-CoV-2 pandemics in a globally connected planet, Math Models Methods Appl Sci, (2024). https://doi.org/10.1142/S0218202524500301 |
| [16] |
A. L. Bertozzi, E. Franco, G. Mohler, M. B. Short, D. Sledge, The challenges of modeling and forecasting the spread of COVID-19, Proc. Natl. Acad. Sci., 117 (2020), 16732–16738. https://doi.org/10.1073/pnas.2006520117 doi: 10.1073/pnas.2006520117
|
| [17] | J. Borghans, R. M. Ribeiro, The maths of memory, eLife, 6 (2017), e26754. https://doi.org/10.7554/eLife.26754 |
| [18] |
D. Burini, D. A. Knopoff, Epidemics and society—A Multiscale vision from the small world to the globally interconnected world, Math Models Methods Appl Sci, 34 (2024), 1564–1594. https://doi.org/10.1142/S0218202524500295 doi: 10.1142/S0218202524500295
|
| [19] |
J. D. Challenger, C. Y. Foo, Y. Wu, A. W. C. Yan, M. M. Marjaneh, F. Liew, et al., Modelling upper respiratory viral load dynamics of SARS-CoV-2, BMC Med, 20 (2022), 25. https://doi.org/10.1186/s12916-021-02220-0 doi: 10.1186/s12916-021-02220-0
|
| [20] |
R. J. De Boer, D. Homann, A. S. Perelson, Different dynamics of CD4$^{+}$ and CD8$^{+}$ T cell responses during and after acute lymphocytic choriomeningitis virus infection, J Immunol, 171 (2003), 3928–3935. https://doi.org/10.4049/jimmunol.171.8.3928 doi: 10.4049/jimmunol.171.8.3928
|
| [21] |
J. Demongeot, Q. Griette, P. Magal, G. Webb, Vaccine efficacy for COVID-19 outbreak in New York City, Biology, 11 (2022), 345. https://doi.org/10.3390/biology11030345 doi: 10.3390/biology11030345
|
| [22] |
M. S. Diamond, T. D. Kanneganti, Innate immunity: the first line of defense against SARS-CoV-2, Nat Immunol, 23 (2022), 165–176. https://doi.org/10.1038/s41590-021-01091-0 doi: 10.1038/s41590-021-01091-0
|
| [23] |
G. Dosi, L. Fanti, M. E. Virgillito, Unequal societies in usual times, unjust societies in pandemic ones, J. Ind. Bus. Econ., 47 (2020), 371–389. https://doi.org/10.1007/s40812-020-00173-8 doi: 10.1007/s40812-020-00173-8
|
| [24] |
R. Eftimie, Grand challenges in mathematical biology: Integrating multi-scale modeling and data, Front Ecol Environ, 8 (2022), 1010622. https://doi.org/10.3389/fams.2022.1010622 doi: 10.3389/fams.2022.1010622
|
| [25] |
I. Eizenberg-Magar, I. Rimer, I. Zaretsky, N. Friedman, Diverse continuum of CD4$^{+}$ T-cell states is determined by hierarchical additive integration of cytokine signals, Proc. Natl. Acad. Sci., 114 (2017), E6447–E6456. https://doi.org/10.1073/pnas.1615590114 doi: 10.1073/pnas.1615590114
|
| [26] |
S. El Zein, O. Chehab, A. Kanj, S. Akrawe, S. Alkassis, T. Mishra et al., SARS-CoV-2 infection: Initial viral load (iVL) predicts severity of illness/outcome, and declining trend of iVL in hospitalized patients corresponds with slowing of the pandemic, PLoS One, 16 (2021), e0255981. https://doi.org/10.1371/journal.pone.0255981 doi: 10.1371/journal.pone.0255981
|
| [27] | M. Elemans, N. K. S. Al Basatena, B. Asquith, The efficiency of the human CD8+ T cell response: how should we quantify it, what determines it, and does it matter? Plos Comput Biol, 8 (2012), e1002381. https://doi.org/10.1371/journal.pcbi.1002381 |
| [28] |
F. Flandoli, E. La Fauci, M. Riva, Individual-based Markov model of virus diffusion: Comparison with COVID-19 incubation period, serial interval and regional time series, Math Models Methods Appl Sci, 31 (2021), 907–939. https://doi.org/10.1142/S0218202521500226 doi: 10.1142/S0218202521500226
|
| [29] | J. F. Fontanari, A stochastic model for the influence of social distancing on loneliness, Physica A, 584 (2021), 126367. |
| [30] |
M. Gatto, E. Bertuzzo, L. Mari, S. Miccoli, L. Carraro, R. Casagrandi, et al., Spread and dynamics of the COVID-19 epidemic in Italy: Effects of emergency containment measures, Proc. Natl. Acad. Sci., 117 (2020), 10484–10491. https://doi.org/10.1073/pnas.2004978117 doi: 10.1073/pnas.2004978117
|
| [31] |
N. M. Gerhards, J. B. W. J. Cornelissen, L. J. M. van Keulen, J. Harders-Westerveen, R. Vloet, B. Smid, et. al., Predictive value of precision-cut lung slices for the susceptibility of three animal species for SARS-CoV-2 and validation in a refined hamster model, Pathogens, 10 (2021), 824. https://doi.org/10.3390/pathogens10070824 doi: 10.3390/pathogens10070824
|
| [32] |
G. Gessain, C. Blériot, F. Ginhoux, Non-genetic heterogeneity of macrophages in diseases–a medical perspective, Front. Cell. Dev. Biol., 8 (2020), 613116. https://doi.org/10.3389/fcell.2020.613116 doi: 10.3389/fcell.2020.613116
|
| [33] |
C. Franceschi, S. Salvioli, P. Garagnani, M de Eguileor, D. Monti, M.Capri, Immunobiography and the heterogeneity of immune responses in the elderly: a focus on inflammaging and trained immunity, Front. Immunol., 8 (2017), 982. https://doi.org/10.3389/fimmu.2017.00982 doi: 10.3389/fimmu.2017.00982
|
| [34] |
J. F. Gianlupi, T. Mapder, T. J. Sego, J. P. Sluka, S. K. Quinney, M. Craig, et al., Multiscale model of antiviral timing, potency, and heterogeneity effects on an epithelial tissue patch infected by SARS-CoV-2, Viruses, 14 (2022), 605. https://doi.org/10.3390/v14030605 doi: 10.3390/v14030605
|
| [35] |
C. H. Hansen, D. Michlmayr, S. M. Gubbels, K. Mølbak, S. Ethelberg, Assessment of protection against reinfection with SARS-CoV-2 among 4 million PCR-tested individuals in Denmark in 2020: a population-level observational study, Lancet, 397 (2021), 1204–1212. https://doi.org/10.1016/S0140-6736(21)00575-4 doi: 10.1016/S0140-6736(21)00575-4
|
| [36] |
P. Hardy, L. S. Marcolino, J. F. Fontanari, The paradox of productivity during quarantine: an agent-based simulation, Eur. Phys. J. B., 94 (2021), 40. https://doi.org/10.1140/epjb/s10051-020-00016-4 doi: 10.1140/epjb/s10051-020-00016-4
|
| [37] |
S. Karimzadeh, R. Bophal, H. N. Tien, Review of infective dose, routes of transmission and outcome of COVID-19 caused by the SARS-CoV-2: comparison with other respiratory viruses, Epidemiol. Infect., 149 (2021), e96. https://doi.org/10.1017/S0950268821000790 doi: 10.1017/S0950268821000790
|
| [38] |
R. Karki, B. R. Sharma, S. Tuladhar, E. P. Williams, L. Zalduondo, P. Samir, et al., Synergism of TNF-$\alpha$ and IFN-$\gamma$ triggers inflammatory cell death, tissue damage, and mortality in SARS-CoV-2 infection and cytokine shock syndrome, Cell, 184 (2021), 149–168. https://doi.org/10.1016/j.cell.2020.11.025 doi: 10.1016/j.cell.2020.11.025
|
| [39] |
D. Kim, A. Quaini, Coupling kinetic theory approaches for pedestrian dynamics and disease contagion in a confined environment, Math Models Methods Appl Sci, 30 (2020), 1893–1915. https://doi.org/10.1142/S0218202520400126 doi: 10.1142/S0218202520400126
|
| [40] |
S. M. Kissler, C. Tedijanto, E. Goldstein, Y. H. Grad, M. Lipsitch, Projecting the transmission dynamics of SARS-CoV-2 through the postpandemic period, Science, 368 (2020), 860–868. https://doi.org/10.1126/science.abb5793 doi: 10.1126/science.abb5793
|
| [41] | Z. Liu, P. Magal, O. Seydi, G. Webb, A model to predict COVID-19 epidemics with applications to South Korea, Italy, and Spain, SIAM News (2020). Available from: https://sinews.siam.org/Details-Page/a-model-to-predict-covid-19-epidemics-with-applications-to-south-korea-italy-and-spain |
| [42] |
S. M. Lynch, G. Guo, D. S. Gibson, A. J. Bjourson, T. Singh Ra, Role of Senescence and Aging in SARS-CoV-2 Infection and COVID-19 Disease, Cells, 10 (2021), 3367–3372. https://doi.org/10.3390/cells10123367 doi: 10.3390/cells10123367
|
| [43] |
D. C. Macallan, R. Busch, B. Asquith, Current estimates of T cell kinetics in humans, Curr. Opin. Syst. Biol., 18 (2019), 77–86. https://doi.org/10.1016/j.coisb.2019.10.002 doi: 10.1016/j.coisb.2019.10.002
|
| [44] |
A. Mantovani, M. Rescigno, G. Forni, F. Tognon, G. Putoto, J. Ictho, P. Lochoro, COVID-19 vaccines and a perspective on Africa, Trends Immunol, 44 (2023), 172–187. https://doi.org/10.1016/j.it.2023.01.005 doi: 10.1016/j.it.2023.01.005
|
| [45] |
J. S. Marshall, R. Warrington, W. Watson, H. L. Kim, An introduction to immunology and immunopathology, Allergy Asthma CL IM, 14 (2018), 49. https://doi.org/10.1186/s13223-018-0289-y doi: 10.1186/s13223-018-0289-y
|
| [46] |
M. Massard, R. Eftimie, A. Perasso, B. Saussereau, A multi-strain epidemic model for COVID-19 with infected and asymptomatic cases: application to French data, J. Theor. Biol., 545 (2022), 111117. https://doi.org/10.1016/j.jtbi.2022.111117 doi: 10.1016/j.jtbi.2022.111117
|
| [47] |
P. M. Matricardi, R. W. Dal Negro, R. Nisini, The first, holistic immunological model of COVID-19: Implications for prevention, diagnosis, and public health measures, Pediatr Allergy Immunol, 31 (2020), 454–470. https://doi.org/10.1111/pai.13271 doi: 10.1111/pai.13271
|
| [48] |
E. Meffre, A. Iwasaki, Interferon deficiency can lead to severe COVID, Nature, 587 (2020), 374–376. https://doi.org/10.1038/d41586-020-03070-1 doi: 10.1038/d41586-020-03070-1
|
| [49] |
P. Moss, The T cell immune response against SARS-CoV-2, Nat. Immunol., 23 (2022), 186–193. https://doi.org/10.1038/s41590-021-01122-w doi: 10.1038/s41590-021-01122-w
|
| [50] | P. Musiani, G. Forni, Basic Immunology, Padua: Piccin, 2018. |
| [51] |
M. G. Netea, J. Dominguez-Andrés, L. B. Barreiro, T. Chavakis, M. Divangahi, E. Fuchs, et al., Defining trained immunity and its role in health and disease, Nat. Rev. Immunol., 20 (2020), 375–388. https://doi.org/10.1038/s41577-020-0285-6 doi: 10.1038/s41577-020-0285-6
|
| [52] |
A. H. Newton, A. Cardani, T. J. Braciale, The host immune response in respiratory virus infection: balancing virus clearance and immunopathology, Semin. Immunol., 38 (2016), 471–482. https://doi.org/10.1007/s00281-016-0558-0 doi: 10.1007/s00281-016-0558-0
|
| [53] |
J. Niessl, T. Sekine, M. Buggert, T cell immunity to SARS-CoV-2, Seminars in Immunology, 55 (2021), 101505. https://doi.org/10.1016/j.smim.2021.101505 doi: 10.1016/j.smim.2021.101505
|
| [54] |
A. Paolini, R. Borella, S. De Biasi, A. Neroni, M. Mattioli, D. Lo Tartaro, et al., Cell death in coronavirus infections: uncovering its role during COVID-19, Cells, 10 (2021), 1585. https://doi.org/10.3390/cells10071585 doi: 10.3390/cells10071585
|
| [55] | B. Perthame, Transport Equations in Biology, Boston: Birkhäuser Basel, 2006. |
| [56] |
J. J. Pinney, F. Rivera-Escalera, C. C. Chu, H. E. Whitehead, K.R. VanDerMeid, A.M. Nelson, et al., Macrophage hypophagia as a mechanism of innate immune exhaustion in mAb-induced cell clearance, Blood, 136 (2020), 2065–2079. https://doi.org/10.1182/blood.2020005571 doi: 10.1182/blood.2020005571
|
| [57] |
D. Pople, E. J. M. Monk, S. Evans, S. Foulkes, J. Islam, E. Wllington, et al., Burden of SARS-CoV-2 infection in healthcare workers during second wave in England and impact of vaccines: prospective multicentre cohort study (SIREN) and mathematical model, BMJ, 378 (2022), e070379. https://doi.org/10.1136/bmj-2022-070379 doi: 10.1136/bmj-2022-070379
|
| [58] |
M. Renardy, C. Hult, S. Evans, J. J. Linderman, D. E. Kirschner, Global sensitivity analysis of biological multiscale models, Curr. Opin. Biomed. Eng., 11 (2019), 109–116. https://doi.org/10.1016/j.cobme.2019.09.012 doi: 10.1016/j.cobme.2019.09.012
|
| [59] |
D. Ricci, M. P. Etna, F. Rizzo, S. Sandini, M. Severa, E. M. Coccia, Innate immune response to SARS-CoV-2 infection: from cells to soluble mediators, Int. J. Mol. Sci., 22 (2021), 7017. https://doi.org/10.3390/ijms22137017 doi: 10.3390/ijms22137017
|
| [60] |
R. Robinot, M. Hubert, G. Dias de Melo, F. Lazarini, T. Bruel, N. Smith, et al., SARS-CoV-2 infection induces the dedifferentiation of multiciliated cells and impairs mucociliary clearance, Nat Commun, 12 (2021), 4354. https://doi.org/10.1038/s41467-021-24521-x doi: 10.1038/s41467-021-24521-x
|
| [61] | Royal Society (Coordinator), RAMP: A call for assistance, addressed to the scientific modelling community. Coordinated by Mark Chaplain, 2021. Available from: https://epcced.github.io/ramp/ |
| [62] |
G. Seminara, B. Carli, G. Forni, S. Fuzzi, A. Mazzino, A. Rinaldo, Biological fluid dynamics of airborne COVID.19 infection, Rend. Fis. Acc. Lincei, 31 (2020), 505–537. https://doi.org/10.1007/s12210-020-00938-2 doi: 10.1007/s12210-020-00938-2
|
| [63] |
A. Seller, C. Hackenbruch, J. S. Walz, A. Nelde, J. S. Heitmann, Long-term follow-up of COVID-19 convalescents–immune response associated with reinfection rate and symptoms, Viruses, 15 (2023), 2100. https://doi.org/10.3390/v15102100 doi: 10.3390/v15102100
|
| [64] |
R. Sender, Y. M. Bar-On, S. Gleizer, B. Bernshtein, A. Flamholz, R. Phillips, et al., The total number and mass of SARS-CoV-2 virions, Proc. Natl. Acad. Sci., 118 (2021), e2024815118. https://doi.org/10.1073/pnas.2024815118 doi: 10.1073/pnas.2024815118
|
| [65] |
A. Sette, S. Crotty, Adaptive immunity to SARS-CoV-2 and COVID-19, Cell, 184 (2021), 861–880. https://doi.org/10.1016/j.cell.2021.01.007 doi: 10.1016/j.cell.2021.01.007
|
| [66] |
H. Shen, D. Chen, C. Li, T. Huang, W. Ma, A mini review of reinfection with the SARS-CoV-2 Omicron variant, Health Sci. Rep., 7 (2024), e2016. https://doi.org/10.1002/hsr2.2016 doi: 10.1002/hsr2.2016
|
| [67] |
A. T. Tan, M. Linster, C. W. Tan, N. L. Bert, W. N. Chia, K. Kunasegaran, et al., Early induction of functional SARS-CoV-2-specific T cells associates with rapid viral clearance and mild disease in COVID-19 patients, Cell Rep., 34 (2021), 108728. https://doi.org/10.1016/j.celrep.2021.108728 doi: 10.1016/j.celrep.2021.108728
|
| [68] |
G. Toscani, P. Sen, S. Biswas, Kinetic exchange models of societies and economies, Philos. Trans. Royal Soc. A, 380 (2022), 20210170. https://doi.org/10.1098/rsta.2021.0170 doi: 10.1098/rsta.2021.0170
|
| [69] | The University of Edinburgh, Review: what is the infectious dose of SARS-CoV-2? Usher Institute, (2021). Available from: https://www.ed.ac.uk/files/atoms/files/uncover_029-01_review_infectious_dose_of_covid-19.pdf. |
| [70] |
J. D. Van Belleghem, P. L. Bollyky, Macrophages and innate immune memory against Staphylococcus skin infections, Proc. Natl. Acad. Sci., 115 (2018), 11865–11867. https://doi.org/10.1073/pnas.1816935115 doi: 10.1073/pnas.1816935115
|
| [71] |
E. Vazquez-Alejo, L. Tarancon-Diez, M. de la Sierra Espinar-Buitrago, M. Genebat, A. Calderón, G. Pérez-Cabeza, et al., Persistent exhausted T-cell immunity after severe COVID-19: 6-month evaluation in a prospective observational study, J. Clin. Med., 12 (2023), 3539. https://doi.org/10.3390/jcm12103539 doi: 10.3390/jcm12103539
|
| [72] |
D. J. Verdon, M. Mulazzani, M.R. Jenkins, Cellular and molecular mechanisms of CD8$^{+}$ T cell differentiation, dysfunction and exhaustion, Int. J. Mol. Sci., 21 (2020), 7357. https://doi.org/10.3390/ijms21197357 doi: 10.3390/ijms21197357
|
| [73] | W. Van Damme, R. Dahake, R. van de Pas, G. Vanham, Y. Assefa, COVID-19: Does the infectious inoculum dose-response relationship contribute to understanding heterogeneity in disease severity and transmission dynamics? Med. Hypotheses, 146 (2021), 110431. https://doi.org/10.1016/j.mehy.2020.110431 |
| [74] |
S. Wang, M. Hao, Z. Pan, J. Lei, X. Zou, Data-driven multiscale mathematical modeling of SARS-CoV-2 infection revels heterogeneity among COVID-19 patients, PLoS Comput. Biol., 17 (2021), e1009587. https://doi.org/10.1371/journal.pcbi.1009587 doi: 10.1371/journal.pcbi.1009587
|
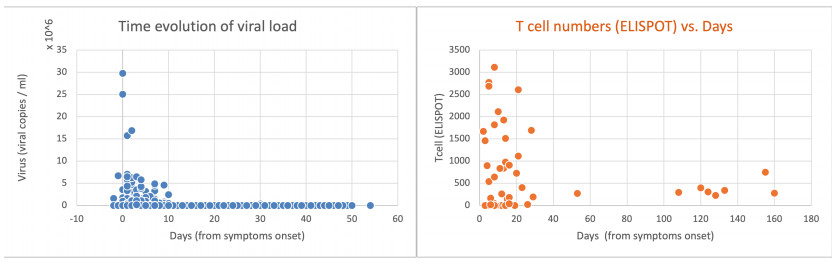







Figures(8) / Tables(2)
Nicola Bellomo, Raluca Eftimie, Guido Forni. What is the in-host dynamics of the SARS-CoV-2 virus? A challenge within a multiscale vision of living systems[J]. Networks and Heterogeneous Media, 2024, 19(2): 655-681. doi: 10.3934/nhm.2024029







 DownLoad:
DownLoad: