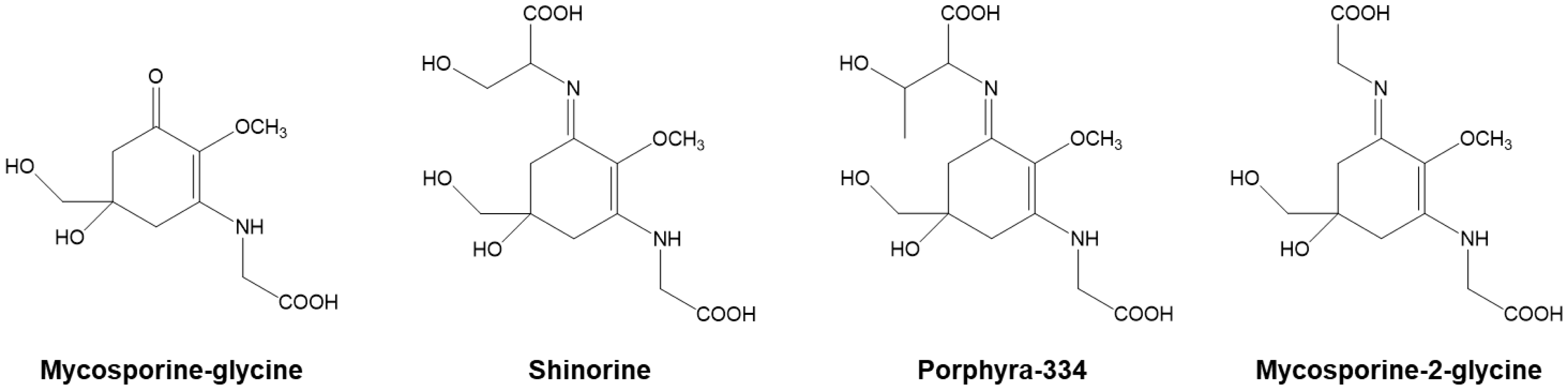
Cyanobacteria are distributed in diverse environments on Earth. They have evolved to synthesize a variety of secondary metabolites to combat and serve as barriers to protect them from distinct environments. Some of these metabolites and compounds exhibit useful physiological activities and are expected to be applied to a wide range of fields, including medicine, agriculture, food, and the cosmetics industry. Due to their photosynthetic ability, cyanobacteria are promising sources for sustainable production of these useful substances. It is expected that future research will lead to the development and efficient production of new compounds. These resources may provide great benefits to our lives and to the environment. This editorial provides an overview of the ultraviolet (UV)-absorbing substances produced by cyanobacteria and discusses future prospects.
Citation: Hakuto Kageyama, Rungaroon Waditee-Sirisattha. Bioactive ultraviolet-absorbing compounds derived from cyanobacteria and microalgae[J]. AIMS Molecular Science, 2025, 12(1): 26-31. doi: 10.3934/molsci.2025002
Cyanobacteria are distributed in diverse environments on Earth. They have evolved to synthesize a variety of secondary metabolites to combat and serve as barriers to protect them from distinct environments. Some of these metabolites and compounds exhibit useful physiological activities and are expected to be applied to a wide range of fields, including medicine, agriculture, food, and the cosmetics industry. Due to their photosynthetic ability, cyanobacteria are promising sources for sustainable production of these useful substances. It is expected that future research will lead to the development and efficient production of new compounds. These resources may provide great benefits to our lives and to the environment. This editorial provides an overview of the ultraviolet (UV)-absorbing substances produced by cyanobacteria and discusses future prospects.

| [1] |
Kageyama H, Waditee-Sirisattha R (2018) Mycosporine-like amino acids as multifunctional secondary metabolites in cyanobacteria: From biochemical to application aspects. Studies in natural products chemistry . Amsterdam, Netherlands: Elsevier 153-194. https://doi.org/10.1016/B978-0-444-64179-3.00005-0 
|
| [2] |
Balskus EP, Walsh CT (2010) The genetic and molecular basis for sunscreen biosynthesis in cyanobacteria. Science 329: 1653-1656. https://doi.org/10.1126/science.1193637
|
| [3] |
Kageyama H, Waditee-Sirisattha R (2022) Mycosporine-like amino acids and scytonemin: Regulations, roles for adaptation of cyanobacteria to the environment, and potential applications. Cyanobacterial physiology: From fundamentals to biotechnology . SanDiego, CA, USA: Academic Press (An imprint of Elsevier) 101-111. https://doi.org/10.1016/B978-0-323-96106-6.00016-2
|
| [4] |
Kageyama H, Waditee-Sirisattha R (2019) Antioxidative, anti-inflammatory, and anti-aging properties of mycosporine-like amino acids: molecular and cellular mechanisms in the protection of skin-aging. Mar Drugs 17: 222. https://doi.org/10.3390/md17040222
|
| [5] |
de la Coba F, Aguilera J, Figueroa FL, et al. (2009) Antioxidant activity of mycosporine-like amino acids isolated from three red macroalgae and one marine lichen. J Appl Phycol 21: 161-169. https://doi.org/10.1007/s10811-008-9345-1
|
| [6] | Jin H, Kim S, Lee D, et al. (2023) Efficient production of mycosporine-like amino acids, natural sunscreens, in Yarrowia lipolytica. Biotechnol Biof Biop 16: 162. https://doi.org/10.1186/s13068-023-02415-y |
| [7] | Park BR, Jeong CR, Cha M, et al. (2024) Sustainable production of shinorine from agricultural wastes using engineered Saccharomyces cerevisiae expressing novel d-alanine-d-alanine ligase from Pseudonocardia pini. J Agric Food Chem 72: 22710-22721. https://doi.org/10.1021/acs.jafc.4c05664 |
| [8] |
Kageyama H, Waditee-Sirisattha R (2023) Distribution, biosynthetic regulation, and bioactivities of mycosporine-2-glycine, a rare UV-protective mycosporine-like amino acid. AIMS Mol Sci 10: 295-310. https://doi.org/10.3934/molsci.2023017
|
| [9] |
Kim S, Park BG, Jin H, et al. (2023) Efficient production of natural sunscreens shinorine, porphyra-334, and mycosporine-2-glycine in Saccharomyces cerevisiae. Metab Eng 78: 137-147. https://doi.org/10.1016/j.ymben.2023.05.009
|
| [10] |
Figueroa FL, Castro-Varela P, Vega J, et al. (2024) Novel synthetic UV screen compounds inspired in mycosporine-like amino acids (MAAs): Antioxidant capacity, photoprotective properties and toxicity. J Photoch Photobiol B 261: 113050. https://doi.org/10.1016/j.jphotobiol.2024.113050
|
| [11] |
Garcia-Pichel F, Castenholz RW (1991) Characterization and biological implications of scytonemin, a cyanobacterial sheath pigment. J Phycol 27: 395-409. https://doi.org/10.1111/j.0022-3646.1991.00395.x
|
| [12] |
Sinha RP, Klisch M, Gröniger A, et al. (2001) Responses of aquatic algae and cyanobacteria to solar UV-B. Plant Ecology 154: 219-236. https://doi.org/10.1023/A:1012986500543
|
| [13] |
Proteau PJ, Gerwick WH, Garcia-Pichel F, et al. (1993) The structure of scytonemin, an ultraviolet sunscreen pigment from the sheaths of cyanobacteria. Experientia 49: 825-829. https://doi.org/10.1007/BF01923559
|
| [14] | Kageyama H, Waditee-Sirisattha R (2018) Cyanobacterial UV sunscreen: Biosynthesis, regulation, and application. Sunscreens: source, formulations, efficacy and recommendations . NOVA Science Publishers 1-28. |
| [15] |
Wada N, Sakamoto T, Matsugo S (2013) Multiple roles of photosynthetic and sunscreen pigments in cyanobacteria focusing on the oxidative stress. Metabolites 3: 463-483. https://doi.org/10.3390/metabo3020463
|
| [16] |
Uchida Y, Maoka T, Palaga T, et al. (2023) Identification of desiccation stress-inducible antioxidative and antiglycative ultraviolet-absorbing oxylipins, saclipin A and saclipin B, in an edible cyanobacterium Aphanothece sacrum. J Agric Food Chem 71: 16137-16147. https://doi.org/10.1021/acs.jafc.3c05152
|
| [17] |
Gerwick WH, Moghaddam M, Hamberg M (1991) Oxylipin metabolism in the red alga Gracilariopsis lemaneiformis: Mechanism of formation of vicinal dihydroxy fatty acids. Arch Biochem Biophys 290: 436-444. https://doi.org/10.1016/0003-9861(91)90563-x
|
| [18] |
Uchida Y, Honda M, Waditee-Sirisattha R, et al. (2024) Photo- and thermo-chemical properties and biological activities of saclipins, UV-absorbing compounds derived from the cyanobacterium Aphanothece sacrum. ACS Agric Sci Technol 4: 1260-1270. https://doi.org/10.1021/acsagscitech.4c00571
|


Figures(3)

Hakuto Kageyama, Rungaroon Waditee-Sirisattha. Bioactive ultraviolet-absorbing compounds derived from cyanobacteria and microalgae[J]. AIMS Molecular Science, 2025, 12(1): 26-31. doi: 10.3934/molsci.2025002







 DownLoad:
DownLoad: