The clinical variability in the course of neuroblastoma (NB) is closely linked to diverse genetic changes acquired by tumor cells. Rapid NB progression is associated with oncogene MYCN amplification (MNA) and segmental chromosomal aberrations (SCA). Alternatively, numerical chromosomal alterations (NCA) have positive impact on treatment. So far, no studies have been undertaken to identify NCA that may group NB patients. Therefore, the aim of the study was to identify NCA typical for NB.
Copy number alterations in NB tumor genome (fresh samples N = 94; formalin-fixed paraffin-embedded specimens N = 66) were analyzed with a pangenomic array CGH technique.
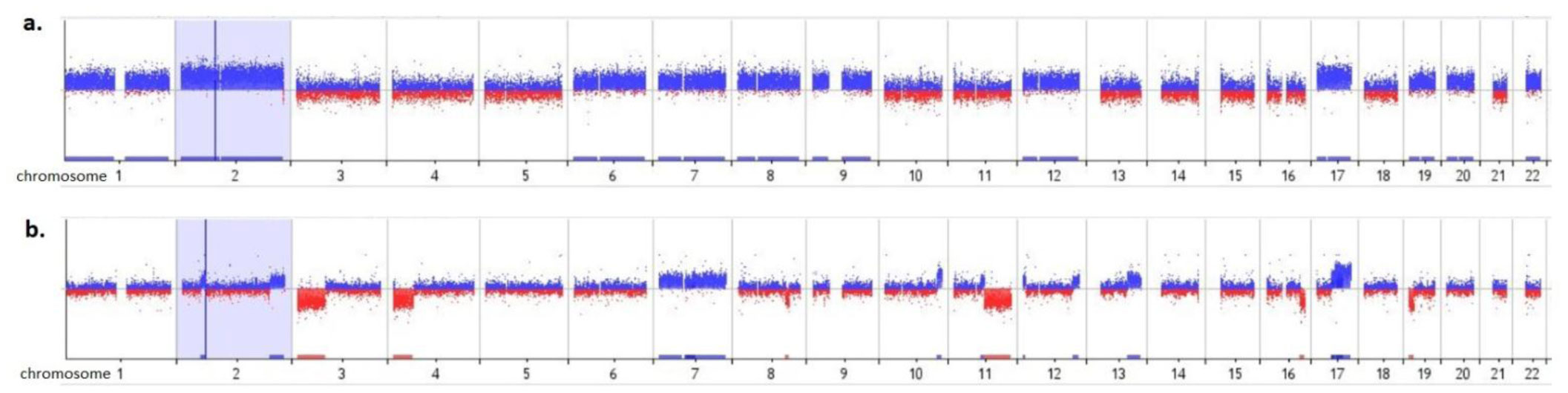
The profile with NCA was observed in 72 (45%) cases, NCA+SCA in 37 (23%), normal in 35 (22%) and MNA in 16 (10%). Samples with NCA were characterized by whole chromosome gains: 17, 7, 6 (78%, 65%, 51%, respectively) and copy loss of chromosome 14 (57%). Similarly to NCA, patients with a combined NCA and SCA profile were also characterized by gain of whole chromosome 17 and 7 (35% both) and loss of chromosome 14 (38%), but with lower frequency. In the combined NCA and SCA profiles, typical NB changes such as deletion 1p36 (27%) and gain 17q (41%) were observed, as well as deletion 11q (24%). The same alterations were detected in MNA samples (44%, 44%, 19%, respectively). A difference was found in spanning 11q deletion between MNA and NCA+SCA subgroup, which may suggest new prognostic markers in NB. In MNA subgroup specific NCA was not indicated.
The hypothesis that NCA in NB tumors are more frequent in younger children with good prognosis was confirmed. To gain new insights into the pathogenesis of NB and to establish molecular targets for diagnosis and therapy, candidate genes in the altered chromosomal regions must be investigated.
Citation: Katarzyna Szewczyk. Typical numerical alterations in genome identified by array CGH analysis in neuroblastoma tumors[J]. AIMS Molecular Science, 2021, 8(4): 248-256. doi: 10.3934/molsci.2021019
The clinical variability in the course of neuroblastoma (NB) is closely linked to diverse genetic changes acquired by tumor cells. Rapid NB progression is associated with oncogene MYCN amplification (MNA) and segmental chromosomal aberrations (SCA). Alternatively, numerical chromosomal alterations (NCA) have positive impact on treatment. So far, no studies have been undertaken to identify NCA that may group NB patients. Therefore, the aim of the study was to identify NCA typical for NB.
Copy number alterations in NB tumor genome (fresh samples N = 94; formalin-fixed paraffin-embedded specimens N = 66) were analyzed with a pangenomic array CGH technique.
The profile with NCA was observed in 72 (45%) cases, NCA+SCA in 37 (23%), normal in 35 (22%) and MNA in 16 (10%). Samples with NCA were characterized by whole chromosome gains: 17, 7, 6 (78%, 65%, 51%, respectively) and copy loss of chromosome 14 (57%). Similarly to NCA, patients with a combined NCA and SCA profile were also characterized by gain of whole chromosome 17 and 7 (35% both) and loss of chromosome 14 (38%), but with lower frequency. In the combined NCA and SCA profiles, typical NB changes such as deletion 1p36 (27%) and gain 17q (41%) were observed, as well as deletion 11q (24%). The same alterations were detected in MNA samples (44%, 44%, 19%, respectively). A difference was found in spanning 11q deletion between MNA and NCA+SCA subgroup, which may suggest new prognostic markers in NB. In MNA subgroup specific NCA was not indicated.
The hypothesis that NCA in NB tumors are more frequent in younger children with good prognosis was confirmed. To gain new insights into the pathogenesis of NB and to establish molecular targets for diagnosis and therapy, candidate genes in the altered chromosomal regions must be investigated.

| [1] |
Mlakar V, Jurkovic-Mlakar S, Lopez G, et al. (2017) 11q deletion in neuroblastoma: a review of biological and clinical implications. Mol Cancer 16: 114-125. doi: 10.1186/s12943-017-0686-8

|
| [2] |
Maris JM, Hogarty MD, Bagatell R, et al. (2007) Neuroblastoma. Lancet 369: 2106-2120. doi: 10.1016/S0140-6736(07)60983-0
|
| [3] |
Cohn SL, Pearson AD, London WB, et al. (2009) The international Neuroblastoma risk group (INRG) classification system: an INRG task force report. J Clin Oncol 27: 289-297. doi: 10.1200/JCO.2008.16.6785
|
| [4] |
Ambros PF, Ambros IM, Brodeur GM, et al. (2009) International consensus for neuroblastoma molecular diagnostics: Report from the International Neuroblastoma Risk Group (INRG) Biology Committee. Br J Cancer 100: 1471-1482. doi: 10.1038/sj.bjc.6605014
|
| [5] |
Canete A, Gerrard M, Rubie H, et al. (2009) Poor survival for infants with MYCNamprified metastatic neuroblastoma despite intensified treatment: The international society of paediatric oncology european neuroblastoma experience. J Clin Oncol 27: 1014-1019. doi: 10.1200/JCO.2007.14.5839
|
| [6] |
Janoueix-Lerosey I, Schleiermacher G, Michels E, et al. (2009) Overall genomic pattern is a predictor of outcome in neuroblastoma. J Clin Oncol 27: 1026-1033. doi: 10.1200/JCO.2008.16.0630
|
| [7] |
Normand C, Michon J, Janoueix-Lerosey I, et al. (2011) Genetic alterations in neuroblastoma and their usefulness for clinical management. Bull Cancer 98: 477-488. doi: 10.1684/bdc.2011.1364
|
| [8] |
Molenaar JJ, Koster J, Zwijnenburg DA, et al. (2012) Sequencing of neuroblastoma identifies chromothripsis and defects in neuritogenesis genes. Nature 483: 589-593. doi: 10.1038/nature10910
|
| [9] |
Lee YH, Kim JH, Song GG (2014) Genome-wide pathway analysis in neuroblastoma. Tumor Biol 35: 3471-3485. doi: 10.1007/s13277-013-1459-7
|
| [10] |
Mwenifumbo JC, Marra MA (2013) Cancer genome-sequencing study design. Nat Rev Genet 14: 321-332. doi: 10.1038/nrg3445
|
| [11] |
Pugh TJ (2013) The genetic landscape of high-risk neuroblastoma. Nat Genet 45: 279-284. doi: 10.1038/ng.2529
|
| [12] |
Thorner PS (2014) The molecular genetic profile of neuroblastoma. Diagn Histopathol 20: 76-83. doi: 10.1016/j.mpdhp.2014.01.005
|
| [13] |
Schleiermacher G, Michon J, Huon I, et al. (2007) Chromosomal CGH identifies patients with a higher risk of relapse in neuroblastoma without MYCN amplification. Br J Cancer 97: 238-246. doi: 10.1038/sj.bjc.6603820
|
| [14] |
Schleiermacher G, Michon J, Ribeiro A, et al. (2011) Segmental chromosomal alterations lead to a higher risk of relapse in infants with MYCN-non-amplified localised unresectable/disseminated neuroblastoma (a SIOPEN collaborative study). Br J Cancer 105: 1940-1948. doi: 10.1038/bjc.2011.472
|
| [15] |
Schleiermacher G, Mosseri V, London WB, et al. (2012) Segmental chromosomal alterations have prognostic impact in neuroblastoma: a report from the INRG project. Br J Cancer 107: 1418-1422. doi: 10.1038/bjc.2012.375
|
| [16] |
Cunsolo CL, Bicocchi MP, Petti AR, et al. (2000) Numerical and structural aberrations in advanced neuroblastoma tumours by CGH analysis; survival correlates with chromosome 17 status. Br J Cancer 83: 1295-1300. doi: 10.1054/bjoc.2000.1432
|
| [17] |
Szewczyk K (2017) Characterization of Chromosomal Aberrations in Neuroblastoma Formalin-Fixed Paraffin-Embedded Specimens with Standard ArrayCGH Procedure - Preliminary Experience. Transl Biomed 8: 117-120. doi: 10.21767/2172-0479.100117
|
| [18] | Ambros IM, Brunner C, Abbasi R, et al. (2014) Ultra-High Density SNParray in Neuroblastoma. Mol Diagn Front Oncol 4: 202-218. |
| [19] |
Hero B, Clement N, Øra I, et al. (2018) Genomic Profiles of Neuroblastoma Associated With Opsoclonus Myoclonus Syndrome. J Pediatr Hematol Oncol 40: 93-98. doi: 10.1097/MPH.0000000000000976
|
| [20] |
Iehara T1, Hamazaki M, Sawada T (2002) Cytogenetic analysis of infantile neuroblastomas by comparative genomic hybridization. Cancer Lett 178: 83-89. doi: 10.1016/S0304-3835(01)00816-3
|
| [21] |
Berbegall AP, Villamón E, Tadeo I, et al. (2014) Neuroblastoma after Childhood: Prognostic Relevance of Segmental Chromosome Aberrations, ATRX Protein Status, and Immune Cell Infiltration. Neoplasia 16: 471-480. doi: 10.1016/j.neo.2014.05.012
|
| [22] |
Stallings RL, Howard J, Dunlop A, et al. (2003) Are gains of chromosomal regions 7q and 11p important abnormalities in neuroblastoma? Cancer Genet Cytogenet 140: 133-137. doi: 10.1016/S0165-4608(02)00681-7
|
| [23] |
Theissen J, Oberthuer A, Hombach A, et al. (2014) Chromosome 17/17q gain and unaltered profiles in high resolution array-CGH are prognostically informative in neuroblastoma. Genes Chromosomes Cancer 53: 639-649. doi: 10.1002/gcc.22174
|
| [24] |
Tonini GP (2017) Growth, progression and chromosome instability of Neuroblastoma: a new scenario of tumorigenesis? BMC Cancer 17: 20-25. doi: 10.1186/s12885-016-2986-6
|
| [25] | Fusco P, Esposito MR, Tonini GP (2018) Chromosome instability in neuroblastoma. Oncol Lett 16: 6887-6894. |
| [26] |
Weaver BA, Cleveland DW (2008) The aneuploidy paradox in cell growth and tumorigenesis. Cancer Cell 14: 431-433. doi: 10.1016/j.ccr.2008.11.011
|
| [27] |
Birkbak NJ, Eklund AC, Li Q, et al. (2011) Paradoxical relationship between chromosomal instability and survival outcome in cancer. Cancer Res 71: 3447-3452. doi: 10.1158/0008-5472.CAN-10-3667
|
| [28] |
Zhang W, Yu Y (2011) The Important Molecular Markers on Chromosome 17 and Their Clinical Impact in Breast Cancer. Int J Mol Sci 12: 5672-5683. doi: 10.3390/ijms12095672
|
| [29] | Saito-Ohara F, Imoto I, Inoue J, et al. (2003) PPM1D is a potential target for 17q gain in neuroblastoma. Cancer Res 63: 1876-1883. |
| [30] | CancerIndex, the Guide to Internet Resources for Cancer family of Web sites. The site was last updated on 12 February, 2021 Available from: http://www.cancerindex.org. |
| [31] |
Couceiro P, Sousa V, Alarcão A, et al. (2010) Polysomy and amplification of chromosome 7 defined for EGFR gene in squamous cell carcinoma of the lung together with exons 19 and 21 wild type. Rev Port Pneumol 16: 453-462. doi: 10.1016/S0873-2159(15)30041-6
|
| [32] |
Buckingham LE, Coon JS, Morrison LE, et al. (2007) The prognostic value of chromosome 7 polysomy in non-small cell lung cancer patients treated with gefitinib. J Thorac Oncol 2: 414-422. doi: 10.1097/01.JTO.0000268675.02744.b0
|
| [33] |
Tabernero MD, Espinosa AB, Maíllo A, et al. (2005) Characterization of chromosome 14 abnormalities by interphase in situ hybridization and comparative genomic hybridization in 124 meningiomas: correlation with clinical, histopathologic, and prognostic features. Am J Clin Pathol 123: 744-751. doi: 10.1309/D7U997XD2PHBCQCN
|
| [34] |
Thompson PM, Seifried BA, Kyemba SK, et al. (2001) Loss of heterozygosity for chromosome 14q in neuroblastoma. Med Pediatr Oncol 36: 28-31. doi: 10.1002/1096-911X(20010101)36:1<28::AID-MPO1008>3.0.CO;2-0
|
| [35] |
Srivatsan ES, Ying KL, Seeger RC, et al. (1993) Deletion of chromosome 11 and of 14q sequences in neuroblastoma. Genes Chromosomes Cancer 7: 32-37. doi: 10.1002/gcc.2870070106
|
| [36] |
Bown N (2001) Neuroblastoma tumour genetics: clinical and biological aspects. J Clin Pathol 54: 897-910. doi: 10.1136/jcp.54.12.897
|
| [37] |
Villamón E, Berbegall AP, Piqueras M, et al. (2013) Genetic instability and intratumoral heterogeneity in neuroblastoma with MYCN amplification plus 11q deletion. PLoS One 8: e53740. doi: 10.1371/journal.pone.0053740
|
| [38] | DECIPHER: Database of Chromosomal Imbalance and Phenotype in Humans using Ensembl Resources. Firth HV et al. Am J Hum Genet 84: 524–533 (2009) .Available from: https://www.deciphergenomics.org/. |
| [39] | Shivapurkar N, Sood S, Wistuba II, et al. (1999) Multiple regions of chromosome 4 demonstrating allelic losses in breast carcinomas. Cancer Res 59: 3576-3580. |
| [40] |
Shivapurkar N, Maitra A, Milchgrub S, et al. (2001) Deletions of chromosome 4 occur early during the pathogenesis of colorectal carcinoma. Hum Pathol 32: 169-177. doi: 10.1053/hupa.2001.21560
|
| [41] | Shivapurkar N, Virmani AK, Wistuba II, et al. (1999) Deletions of chromosome 4 at multiple sites are frequent in malignant mesothelioma and small cell lung carcinoma. Clin Cancer Res 5: 17-23. |
| [42] |
Brinkschmidt C, Poremba C, Christiansen H, et al. (1998) Comparative genomic hybridization and telomerase activity analysis identify two biologically different groups of 4s neuroblastomas. Br J Cancer 77: 2223-2229. doi: 10.1038/bjc.1998.370
|
Figures(1) / Tables(1)

Katarzyna Szewczyk. Typical numerical alterations in genome identified by array CGH analysis in neuroblastoma tumors[J]. AIMS Molecular Science, 2021, 8(4): 248-256. doi: 10.3934/molsci.2021019







 DownLoad:
DownLoad: