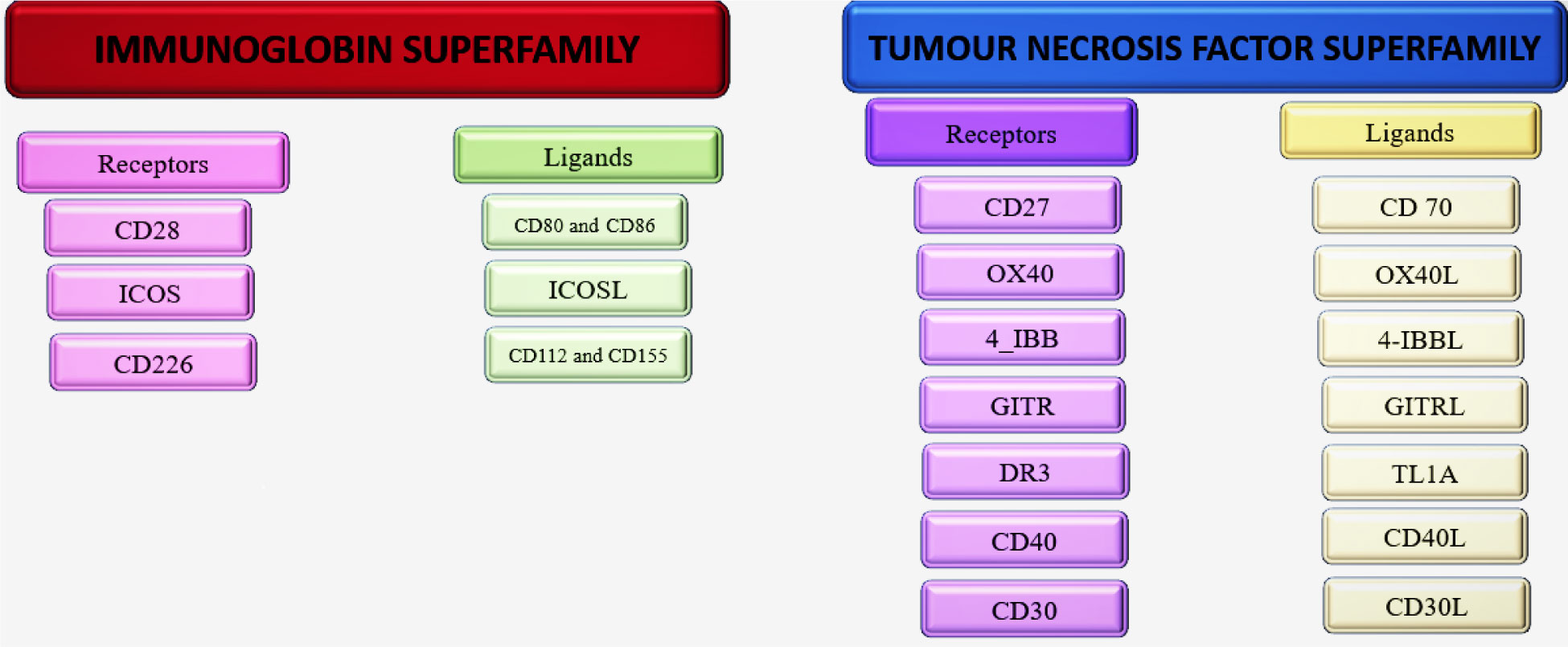
Body's defence mechanism has ability to combat tumour cells but tumour cells can circumvent immune system in order to flourish. Therefore, current research focuses on reinvigorating immune system to combat against extensive range of human malignancies through immunotherapy. Recently, immuno-therapy has demonstrated beneficial outcomes in cancers treatment but the main drawbacks are primary and acquired resistance to the therapeutic agents and immune-related toxicities. Therefore, novel immune therapies are direly required. Co-stimulatory molecules such as TNF Receptor Superfamily Member 4 (OX40, CD134) and its ligand TNF Superfamily Member 4 (CD252, OX40L) are expressed on different immune cells. The mutual interaction between OX40 and its ligand (OX40/OX40L) decreases the functional capacity of immunosuppression offered by regulatory T cells (Tregs) and induces the proliferation of T cells against specific antigen enhancing the immune response. Many clinical trials are focusing on OX40/OX40L therapeutic agents to find out whether they have therapeutic effect on cancer treatment. The initial phase trials result of OX40 and its ligands focusing therapeutic agents are encouraging but still not sufficient. This review will concentrate on the cellular and molecular pathways of OX40-mediated T-cell co-stimulation, the expression of OX40 and OX40L in tumours, the implications of their interactions and their under-or over-expression patterns, with particular focus on the function of OX40 in tumours of different origins. Finally, we discuss results of clinical trials of OX40 and OX40L directed pharmacotherapy and the lacunae that need to be filled.
Citation: Aliya I Sani, Zil-e-Rubab, Shumaila Usman, Syed Zaryab Ahmed, Mervyn Hosein. Role of OX40 and its ligand as costimulatory modulators in cancer immunotherapy[J]. AIMS Molecular Science, 2021, 8(3): 161-173. doi: 10.3934/molsci.2021012
Body's defence mechanism has ability to combat tumour cells but tumour cells can circumvent immune system in order to flourish. Therefore, current research focuses on reinvigorating immune system to combat against extensive range of human malignancies through immunotherapy. Recently, immuno-therapy has demonstrated beneficial outcomes in cancers treatment but the main drawbacks are primary and acquired resistance to the therapeutic agents and immune-related toxicities. Therefore, novel immune therapies are direly required. Co-stimulatory molecules such as TNF Receptor Superfamily Member 4 (OX40, CD134) and its ligand TNF Superfamily Member 4 (CD252, OX40L) are expressed on different immune cells. The mutual interaction between OX40 and its ligand (OX40/OX40L) decreases the functional capacity of immunosuppression offered by regulatory T cells (Tregs) and induces the proliferation of T cells against specific antigen enhancing the immune response. Many clinical trials are focusing on OX40/OX40L therapeutic agents to find out whether they have therapeutic effect on cancer treatment. The initial phase trials result of OX40 and its ligands focusing therapeutic agents are encouraging but still not sufficient. This review will concentrate on the cellular and molecular pathways of OX40-mediated T-cell co-stimulation, the expression of OX40 and OX40L in tumours, the implications of their interactions and their under-or over-expression patterns, with particular focus on the function of OX40 in tumours of different origins. Finally, we discuss results of clinical trials of OX40 and OX40L directed pharmacotherapy and the lacunae that need to be filled.
antigen presenting cells
cytotoxic T-lymphocyte-associated protein 4
forkhead box P3
Hepatocellular carcinoma
major histocompatibility complex
Natural Killer cell
Non-small cell lung carcinoma
Oral squamous cell carcinoma
Tumour necrosis factor receptor superfamily member 4
Tumour necrosis factor superfamily member 4 ligand
T cell receptor
Regulatory T cells

| [1] |
Hanahan D, Weinberg Robert A (2011) Hallmarks of cancer: the next generation. Cell 144: 646-674. doi: 10.1016/j.cell.2011.02.013

|
| [2] |
Zappasodi R, Merghoub T, Wolchok JD (2018) Emerging concepts for immune checkpoint Blockade-based combination therapies. Cancer Cell 34: 690. doi: 10.1016/j.ccell.2018.09.008
|
| [3] |
Farkona S, Diamandis EP, Blasutig IM (2016) Cancer immunotherapy: the beginning of the end of cancer? BMC Med 14: 1-18. doi: 10.1186/s12916-016-0623-5
|
| [4] |
Fares CM, Van Allen EM, Drake CG, et al. (2019) Mechanisms of resistance to immune checkpoint blockade: why does checkpoint inhibitor immunotherapy not work for all patients? Am Soc Clin Oncol Educ Book 39: 147-164. doi: 10.1200/EDBK_240837
|
| [5] |
Croft M (2010) Control of immunity by the TNFR-related molecule OX40 (CD134). Annu Rev Immunol 28: 57-78. doi: 10.1146/annurev-immunol-030409-101243
|
| [6] |
Alves Costa Silva C, Facchinetti F, Routy B, et al. (2020) New pathways in immune stimulation: targeting OX40. ESMO Open 5: e000573. doi: 10.1136/esmoopen-2019-000573
|
| [7] |
Baum PR, Gayle RB, Ramsdell F, et al. (1994) Molecular characterization of murine and human OX40/OX40 ligand systems: identification of a human OX40 ligand as the HTLV-1-regulated protein gp34. EMBO J 13: 3992-4001. doi: 10.1002/j.1460-2075.1994.tb06715.x
|
| [8] |
Croft M, So T, Duan W, et al. (2009) The significance of OX40 and OX40L to T-cell biology and immune disease. Immunol Rev 229: 173-191. doi: 10.1111/j.1600-065X.2009.00766.x
|
| [9] |
Kawamata S, Hori T, Imura A, et al. (1998) Activation of OX40 signal transduction pathways leads to tumor necrosis factor receptor-associated factor (TRAF) 2- and TRAF5-mediated NF-κB activation. J Biol Chem 273: 5808-5814. doi: 10.1074/jbc.273.10.5808
|
| [10] |
Zhang X, Xiao X, Lan P, et al. (2018) OX40 costimulation inhibits Foxp3 expression and Treg induction via BATF3-dependent and independent mechanisms. Cell Rep 24: 607-618. doi: 10.1016/j.celrep.2018.06.052
|
| [11] |
Watts TH (2005) TNF/TNFR family members in co-stimulation of T cell responses. Annu Rev Immunol 23: 23-68. doi: 10.1146/annurev.immunol.23.021704.115839
|
| [12] |
Redmond WL, Ruby CE, Weinberg AD (2009) The role of OX40-mediated co-stimulation in T-cell activation and survival. Crit Rev Immunol 29: 187-201. doi: 10.1615/CritRevImmunol.v29.i3.10
|
| [13] |
Kashiwakura J, Yokoi H, Saito H, et al. (2004) T cell proliferation by direct cross-talk between OX40 ligand on human mast cells and OX40 on human T cells: comparison of gene expression profiles between human tonsillar and lung-cultured mast cells. J Immunol 173: 5247-5257. doi: 10.4049/jimmunol.173.8.5247
|
| [14] |
Turaj AH, Cox KL, Penfold CA, et al. (2018) Augmentation of CD134 (OX40)-dependent NK anti-tumour activity is dependent on antibody cross-linking. Sci Rep 8: 2278. doi: 10.1038/s41598-018-20656-y
|
| [15] |
Ramser M, Eichelberger S, Däster S, et al. (2018) High OX40 expression in recurrent ovarian carcinoma is indicative for response to repeated chemotherapy. BMC Cancer 18: 425. doi: 10.1186/s12885-018-4339-0
|
| [16] | Hamidinia M, Boroujerdnia MG, Talaiezadeh A, et al. (2013) Concomitant increase of OX40 and FOXP3 transcripts in peripheral blood of patients with breast Ccancer. Iran J Immunol 10: 22-30. |
| [17] |
Massarelli E, Lam VK, Parra ER, et al. (2019) High OX-40 expression in the tumor immune infiltrate is a favorable prognostic factor of overall survival in non-small cell lung cancer. J Immunother Cancer 7: 351. doi: 10.1186/s40425-019-0827-2
|
| [18] |
Lecerf C, Kamal M, Vacher S, et al. (2019) Immune gene expression in head and neck squamous cell carcinoma patients. Eur J Cancer 121: 210-223. doi: 10.1016/j.ejca.2019.08.028
|
| [19] |
Martins MR, Santos RLD, Jatahy KDN, et al. (2018) Could OX40 agonist antibody promote activation of the anti-tumor immune response in gastric cancer. J Surg Oncol 117: 840-844. doi: 10.1002/jso.25001
|
| [20] | Sawada R, Arai Y, Sagawa Y, et al. (2019) High blood levels of soluble OX40 (CD134), an immune costimulatory molecule, indicate reduced survival in patients with advanced colorectal cancer. Oncol Rep 42: 2057-2064. |
| [21] |
He Y, Zhang X, Jia K, et al. (2019) OX40 and OX40L protein expression of tumor infiltrating lymphocytes in non-small cell lung cancer and its role in clinical outcome and relationships with other immune biomarkers. Transl Lung Cancer Res 8: 352-366. doi: 10.21037/tlcr.2019.08.15
|
| [22] |
Roszik J, Markovits E, Dobosz P, et al. (2019) TNFSF4 (OX40L) expression and survival in locally advanced and metastatic melanoma. Cancer Immunol Immunother 68: 1493-1500. doi: 10.1007/s00262-019-02382-0
|
| [23] |
Xie K, Xu L, Wu H, et al. (2018) OX40 expression in hepatocellular carcinoma is associated with a distinct immune microenvironment, specific mutation signature, and poor prognosis. Oncoimmunology 7: e1404214. doi: 10.1080/2162402X.2017.1404214
|
| [24] |
Baruah P, Lee M, Odutoye T, et al. (2012) Decreased levels of alternative co-stimulatory receptors OX40 and 4-1BB characterise T cells from head and neck cancer patients. Immunobiology 217: 669-675. doi: 10.1016/j.imbio.2011.11.005
|
| [25] |
Montler R, Bell RB, Thalhofer C, et al. (2016) OX40, PD-1 and CTLA-4 are selectively expressed on tumor-infiltrating T cells in head and neck cancer. Clin Transl Immunology 5: e70. doi: 10.1038/cti.2016.16
|
| [26] | Sani AI, Rubab ZE, Usman S, et al. (2021) Serum levels of OX40 in early and late-stage oral squamous cell carcinoma. Cureus 13: e14597. |
| [27] |
Du P, Wang Z, Geng J, et al. (2021) Expression and clinical significance of OX40 and OX40L mRNA in hepatocellular carcinoma. Bull Exp Biol Med 170: 485-488. doi: 10.1007/s10517-021-05093-8
|
| [28] |
Kashima J, Okuma Y, Hosomi Y, et al. (2020) High serum OX40 and OX40 ligand (OX40L) levels correlate with reduced survival in patients with advanced lung adenocarcinoma. Oncology 98: 303-310. doi: 10.1159/000505975
|
| [29] |
Redmond WL, Linch SN, Kasiewicz MJ (2013) Combined targeting of co-stimulatory (OX40) and co-inhibitory (CTLA-4) pathways elicits potent effector T cells capable of driving robust anti-tumor immunity. Cancer Immunol Res 2: 142-153. doi: 10.1158/2326-6066.CIR-13-0031-T
|
| [30] |
Jahan N, Talat H, Curry WT (2017) Agonist OX40 immunotherapy improves survival in glioma-bearing mice and is complementary with vaccination with irradiated GM-CSF–expressing tumor cells. Neuro Oncol 20: 44-54. doi: 10.1093/neuonc/nox125
|
| [31] |
Duhen R, Ballesteros-Merino C, Frye AK, et al. (2021) Neoadjuvant anti-OX40 (MEDI6469) therapy in patients with head and neck squamous cell carcinoma activates and expands antigen-specific tumor-infiltrating T cells. Nat Commun 12: 1047. doi: 10.1038/s41467-021-21383-1
|
| [32] |
Bell RB, Duhen R, Leidner RS, et al. (2018) Neoadjuvant anti-OX40 (MEDI6469) prior to surgery in head and neck squamous cell carcinoma. J Clin Oncol 36: 6011-6011. doi: 10.1200/JCO.2018.36.15_suppl.6011
|
| [33] |
El-Khoueiry AB, Spano JP, Angevin E, et al. (2020) Analysis of OX40 agonist antibody (PF-04518600) in patients with hepatocellular carcinoma. J Clin Oncol 38: 523-523. doi: 10.1200/JCO.2020.38.4_suppl.523
|
| [34] |
Griffiths J, Hussain K, Smith HL, et al. (2020) Domain binding and isotype dictate the activity of anti-human OX40 antibodies. J Immunother Cancer 28: e001557. doi: 10.1136/jitc-2020-001557
|
| [35] |
Gough MJ, Ruby CE, Redmond WL, et al. (2008) OX40 agonist therapy enhances CD8 infiltration and decreases immune suppression in the tumor. Cancer Res 68: 5206-5215. doi: 10.1158/0008-5472.CAN-07-6484
|
| [36] |
Bantia S, Choradia N (2018) Treatment duration with immune-based therapies in cancer: an enigma. J Immunother Cancer 6: 143. doi: 10.1186/s40425-018-0465-0
|
| [37] |
Kjaergaard J, Peng L, Cohen PA, et al. (2001) Augmentation versus inhibition: effects of conjunctional OX-40 receptor monoclonal antibody and IL-2 treatment on adoptive immunotherapy of advanced tumor. J Immunol 167: 6669-6677. doi: 10.4049/jimmunol.167.11.6669
|
| [38] |
Morris A, Vetto JT, Ramstad T, et al. (2001) Induction of anti-mammary cancer immunity by engaging the OX-40 receptor in vivo. Breast Cancer Res Treat 67: 71-80. doi: 10.1023/A:1010649303056
|
| [39] |
Guo Z, Wang X, Cheng D, et al. (2014) PD-1 blockade and OX40 triggering synergistically protects against tumor growth in a murine model of ovarian cancer. Plos One 9: e89350. doi: 10.1371/journal.pone.0089350
|
| [40] |
Messenheimer DJ, Jensen SM, Afentoulis ME, et al. (2017) Timing of PD-1 blockade is critical to effective combination immunotherapy with anti-OX40. Clin Cancer Res 23: 6165-6177. doi: 10.1158/1078-0432.CCR-16-2677
|
| [41] |
Linch SN, Kasiewicz MJ, McNamara MJ, et al. (2016) Combination OX40 agonism/CTLA-4 blockade with HER2 vaccination reverses T-cell anergy and promotes survival in tumor-bearing mice. Proc Natl Acad Sci U S A 113: E319-E327. doi: 10.1073/pnas.1510518113
|
| [42] |
Kvarnhammar AM, Veitonmäki N, Hägerbrand K, et al. (2019) The CTLA-4 x OX40 bispecific antibody ATOR-1015 induces anti-tumor effects through tumor-directed immune activation. J Immunother Cancer 7: 103. doi: 10.1186/s40425-019-0570-8
|
| [43] |
Andarini S, Kikuchi T, Nukiwa M, et al. (2004) Adenovirus vector-Mediated in vivo gene transfer of OX40 ligand to tumor cells enhances antitumor immunity of tumor-bearing hosts. Cancer Res 64: 3281-3287. doi: 10.1158/0008-5472.CAN-03-3911
|
| [44] |
Sadun RE, Hsu WE, Zhang N, et al. (2008) Fc-mOX40L fusion protein produces complete remission and enhanced survival in 2 murine tumor models. J Immunother 31: 235-245. doi: 10.1097/CJI.0b013e31816a88e0
|
| [45] |
Jahan N, Talat H, Alonso A, et al. (2019) Triple combination immunotherapy with GVAX, anti-PD-1 monoclonal antibody, and agonist anti-OX40 monoclonal antibody is highly effective against murine intracranial glioma. Oncoimmunology 8: e1577108. doi: 10.1080/2162402X.2019.1577108
|
| [46] |
Berrong Z, Mkrtichyan M, Ahmad S, et al. (2018) Antigen-specific antitumor responses induced by OX40 agonist are enhanced by the IDO inhibitor indoximod. Cancer Immunol Res 6: 201-208. doi: 10.1158/2326-6066.CIR-17-0223
|

Figures(2) / Tables(2)

Aliya I Sani, Zil-e-Rubab, Shumaila Usman, Syed Zaryab Ahmed, Mervyn Hosein. Role of OX40 and its ligand as costimulatory modulators in cancer immunotherapy[J]. AIMS Molecular Science, 2021, 8(3): 161-173. doi: 10.3934/molsci.2021012







 DownLoad:
DownLoad: