Inflammation and oxidative stress are implicated in several chronic disorders, while healthy foods and especially fermented beverages and those containing probiotics can provide anti-inflammatory and antioxidant protection against such manifestations and the associated disorders. Water kefir is such a beverage that is rich in both probiotic microbiota and anti-inflammatory bioactives, with an increasing demand as an alternative to a fermented product based on non-dairy matrix with potential health properties. Within this study, the health-promoting properties of the most representative species and strains of microorganisms present in water kefir grains, as well as the health benefits attributed to the bioactive metabolites produced by each individual strain in a series of their cultures, were thoroughly reviewed. Emphasis was given to the antioxidant, antithrombotic, and anti-inflammatory bio-functionalities of both the cultured microorganisms and the bioactive metabolites produced in each case. Moreover, an extensive presentation of the antioxidant and anti-inflammatory health benefits observed from the overall water kefir cultures and classic water kefir beverages obtained were also conducted. Finally, the use of water kefir for the production of several other bio-functional products, including fermented functional foods, supplements, nutraceuticals, nutricosmetics, cosmeceuticals, and cosmetic applications with anti-inflammatory and antioxidant health promoting potential was also thoroughly discussed. Limitations and future perspectives on the use of water kefir, its microorganisms, and their bioactive metabolites are also outlined.
Citation: Dimitra Papadopoulou, Vasiliki Chrysikopoulou, Aikaterini Rampaouni, Alexandros Tsoupras. Antioxidant and anti-inflammatory properties of water kefir microbiota and its bioactive metabolites for health promoting bio-functional products and applications[J]. AIMS Microbiology, 2024, 10(4): 756-811. doi: 10.3934/microbiol.2024034
Inflammation and oxidative stress are implicated in several chronic disorders, while healthy foods and especially fermented beverages and those containing probiotics can provide anti-inflammatory and antioxidant protection against such manifestations and the associated disorders. Water kefir is such a beverage that is rich in both probiotic microbiota and anti-inflammatory bioactives, with an increasing demand as an alternative to a fermented product based on non-dairy matrix with potential health properties. Within this study, the health-promoting properties of the most representative species and strains of microorganisms present in water kefir grains, as well as the health benefits attributed to the bioactive metabolites produced by each individual strain in a series of their cultures, were thoroughly reviewed. Emphasis was given to the antioxidant, antithrombotic, and anti-inflammatory bio-functionalities of both the cultured microorganisms and the bioactive metabolites produced in each case. Moreover, an extensive presentation of the antioxidant and anti-inflammatory health benefits observed from the overall water kefir cultures and classic water kefir beverages obtained were also conducted. Finally, the use of water kefir for the production of several other bio-functional products, including fermented functional foods, supplements, nutraceuticals, nutricosmetics, cosmeceuticals, and cosmetic applications with anti-inflammatory and antioxidant health promoting potential was also thoroughly discussed. Limitations and future perspectives on the use of water kefir, its microorganisms, and their bioactive metabolites are also outlined.

| [1] | World health statistics 2023: monitoring health for the SDGs, sustainable development goals. Available from: https://www.who.int/publications/i/item/9789240074323 |
| [2] | Noncommunicable diseases-World Health Organization (WHO) 2023 (16 September). Available from: https://www.who.int/news-room/fact-sheets/detail/noncommunicable-diseases |
| [3] | Noncommunicable diseases-World Health Organization (WHO) 2023 (Dec 20). Available from: https://www.who.int/health-topics/noncommunicable-diseases#tab=tab_1 |
| [4] |
Boutayeb A, Boutayeb S (2005) The burden of non communicable diseases in developing countries. Int J Equity Health 4: 2. https://doi.org/10.1186/1475-9276-4-2 
|
| [5] | Diabetes Prevention Program Research Group.Reduction in the Incidence of Type 2 Diabetes with Lifestyle Intervention or Metformin. N Engl J Med (2002) 346: 393-403. https://doi.org/10.1056/NEJMoa012512 |
| [6] | Tsoupras A, Iatrou C, Frangia C, et al. (2009) The implication of platelet activating factor in cancer growth and metastasis: potent beneficial role of PAF-inhibitors and antioxidants. Infect Disord-Drug Targets Former Curr Drug Targets-Infect Disord 9: 390-399. https://doi.org/10.2174/187152609788922555 |
| [7] |
Tsoupras A, Lordan R, Zabetakis I (2018) Inflammation, not cholesterol, is a cause of chronic disease. Nutrients 10: 604. https://doi.org/10.3390/nu10050604
|
| [8] |
Tsoupras A, Lordan R, Zabetakis I (2019) Inflammation and cardiovascular diseases. The Impact of Nutrition and Statins on Cardiovascular Diseases . Elsevier 53-117. https://doi.org/10.1016/B978-0-12-813792-5.00003-3
|
| [9] |
Nathan C, Ding A (2010) Nonresolving inflammation. Cell 140: 871-882. https://doi.org/10.1016/j.cell.2010.02.029
|
| [10] |
Peluso I, Morabito G, Urban L, et al. (2012) Oxidative stress in atherosclerosis development: the central role of LDL and oxidative burst. Endocr Metab Immune Disord-Drug Targets 12: 351-360. https://doi.org/10.2174/187153012803832602
|
| [11] |
Valko M, Leibfritz D, Moncol J, et al. (2007) Free radicals and antioxidants in normal physiological functions and human disease. Int J Biochem Cell Biol 39: 44-84. https://doi.org/10.1016/j.biocel.2006.07.001
|
| [12] |
Koloverou E, Panagiotakos DB, Pitsavos C, et al. (2016) Adherence to Mediterranean diet and 10-year incidence (2002–2012) of diabetes: correlations with inflammatory and oxidative stress biomarkers in the ATTICA cohort study. Diabetes Metab Res Rev 32: 73-81. https://doi.org/10.1002/dmrr.2672
|
| [13] |
Estruch R, Martínez-González MA, Corella D, et al. (2006) Effects of a Mediterranean-style diet on cardiovascular risk factors: a randomized trial. Ann Intern Med 145: 1-11. https://doi.org/10.7326/0003-4819-145-1-200607040-00004
|
| [14] | Zabetakis I, Lordan R, Tsoupras A, et al. Functional foods and their implications for health promotion, Academic Press (2022). https://doi.org/10.1016/B978-0-12-823811-0.00005-5 |
| [15] | Tsoupras A, Panagopoulou EA, Kyzas GZ (2024) Anti-inflammatory, antithrombotic and anti-oxidant bioactives of beer and brewery by-products, as ingredients of bio-functional foods, nutraceuticals, cosmetics, cosmeceuticals and pharmaceuticals with health promoting properties. AIMS Agric Food . https://doi.org/10.3934/agrfood.2024032 |
| [16] |
Tsoupras A, Ni VLJ, O'Mahony É, et al. (2023) Winemaking: “with one stone, two birds”? a holistic review of the bio-functional compounds, applications and health benefits of wine and wineries' by-products. Fermentation 9: 838. https://doi.org/10.3390/fermentation9090838
|
| [17] |
Moran D, Fleming M, Daly E, et al. (2021) Anti-platelet properties of apple must/skin yeasts and of their fermented apple cider products. Beverages 7: 54. https://doi.org/10.3390/beverages7030054
|
| [18] | Tsoupras A, Gkika D, Markopoulos T, et al. (2024) Apple products (apple juice and cider) and by-products (apple pomace): Bioactive compounds and biological properties. Springer . https://doi.org/10.1007/978-3-031-04195-2_214-1 |
| [19] |
Conde TA, Zabetakis I, Tsoupras A, et al. (2021) Microalgal lipid extracts have potential to modulate the inflammatory response: a critical review. Int J Mol Sci 22: 9825. https://doi.org/10.3390/ijms22189825
|
| [20] |
Tsoupras A, Davi KG (2024) Bioactive metabolites from fungi with anti-inflammatory and antithrombotic properties: current status and future perspectives for drug development. Fungi Bioactive Metabolites . Singapore: Springer Nature Singapore 427-494. https://doi.org/10.1007/978-981-99-5696-8_14
|
| [21] |
Thiyagarasaiyar K, Goh BH, Jeon YJ, et al. (2020) Algae metabolites in cosmeceutical: an overview of current applications and challenges. Mar Drugs 18: 323. https://doi.org/10.3390/md18060323
|
| [22] |
Chrzanowski G (2020) Saccharomyces Cerevisiae—an interesting producer of bioactive plant polyphenolic metabolites. Int J Mol Sci 21: 7343. https://doi.org/10.3390/ijms21197343
|
| [23] | Culpepper T (2022) The effects of kefir and kefir components on immune and metabolic physiology in pre-clinical studies: a narrative review. Cureus . https://doi.org/10.7759/cureus.27768 |
| [24] |
Apalowo OE, Adegoye GA, Mbogori T, et al. (2024) Nutritional characteristics, health impact, and applications of kefir. Foods 13: 1026. https://doi.org/10.3390/foods13071026
|
| [25] |
Moretti AF, Moure MC, Quiñoy F, et al. (2022) Water kefir, a fermented beverage containing probiotic microorganisms: From ancient and artisanal manufacture to industrialized and regulated commercialization. Future Foods 5: 100123. https://doi.org/10.1016/j.fufo.2022.100123
|
| [26] |
Marsh AJ, O'Sullivan O, Hill C, et al. (2013) Sequence-based analysis of the microbial composition of water kefir from multiple sources. FEMS Microbiol Lett 348: 79-85. https://doi.org/10.1111/1574-6968.12248
|
| [27] |
Romero-Luna HE, Peredo-Lovillo A, Hernández-Mendoza A, et al. (2020) Probiotic potential of Lactobacillus paracasei CT12 isolated from water kefir grains (Tibicos). Curr Microbiol 77: 2584-2592. https://doi.org/10.1007/s00284-020-02016-0
|
| [28] |
Peluzio MDCG, Dias MDME, Martinez JA, et al. (2021) Kefir and intestinal microbiota modulation: implications in human health. Front Nutr 8: 638740. https://doi.org/10.3389/fnut.2021.638740
|
| [29] |
Zavala L, Golowczyc MA, Van Hoorde K, et al. (2016) Selected Lactobacillus strains isolated from sugary and milk kefir reduce Salmonella infection of epithelial cells in vitro. Benef Microbes 7: 585-595. https://doi.org/10.3920/BM2015.0196
|
| [30] |
Zamberi NR, Abu N, Mohamed NE, et al. (2016) The antimetastatic and antiangiogenesis effects of kefir water on murine breast cancer cells. Integr Cancer Ther 15: NP53-NP66. https://doi.org/10.1177/1534735416642862
|
| [31] |
Gamba RR, Yamamoto S, Sasaki T, et al. (2019) Microbiological and functional characterization of kefir grown in different sugar solutions. Food Sci Technol Res 25: 303-312. https://doi.org/10.3136/fstr.25.303
|
| [32] | Koh WY, Utra U, Rosma A, et al. (2017) Development of a novel fermented pumpkin-based beverage inoculated with water kefir grains: a response surface methodology approach. Food Sci Biotechnol . https://doi.org/10.1007/s10068-017-0245-5 |
| [33] |
Diniz RO, Garla LK, Schneedorf JM, et al. (2003) Study of anti-inflammatory activity of Tibetan mushroom, a symbiotic culture of bacteria and fungi encapsulated into a polysaccharide matrix. Pharmacol Res 47: 49-52. https://doi.org/10.1016/S1043-6618(02)00240-2
|
| [34] |
Darvishzadeh P, Orsat V, Martinez JL (2021) Process optimization for development of a novel water kefir drink with high antioxidant activity and potential probiotic properties from russian olive fruit (Elaeagnus angustifolia). Food Bioprocess Technol 14: 248-260. https://doi.org/10.1007/s11947-020-02563-1
|
| [35] | Yusuf D, Nuraida L, Dewanti-Hariyadi R, et al. (2020) In vitro antioxidant and α-Glucosidase inhibitory activities of Lactobacillus spp. isolated from indonesian kefir grains. Appl Food Biotechnol 8: 39-46. https://doi.org/10.22037/afb.v8i1.30367 |
| [36] |
Mäkinen OE, Wanhalinna V, Zannini E, et al. (2016) Foods for special dietary needs: non-dairy plant-based milk substitutes and fermented dairy-type products. Crit Rev Food Sci Nutr 56: 339-349. https://doi.org/10.1080/10408398.2012.761950
|
| [37] |
Fiorda FA, De Melo Pereira GV, Thomaz-Soccol V, et al. (2016) Development of kefir-based probiotic beverages with DNA protection and antioxidant activities using soybean hydrolyzed extract, colostrum and honey. LWT-Food Sci Technol 68: 690-697. https://doi.org/10.1016/j.lwt.2016.01.003
|
| [38] |
Gulitz A, Stadie J, Wenning M, et al. (2011) The microbial diversity of water kefir. Int J Food Microbiol 151: 284-288. https://doi.org/10.1016/j.ijfoodmicro.2011.09.016
|
| [39] | The ferment company Water Kefir Starter. Available from: https://www.thefermentcompany.nl/product/waterkefir-starter/ |
| [40] | Hecer C, Ulusoy B, Kaynarca D (2019) Effect of different fermentation conditions on composition of kefir microbiota. Int Food Res J 26. Available from: https://www.researchgate.net/publication/333561113_Effect_of_different_fermentation_conditions_on_composition_of_kefir_microbiota |
| [41] |
Verce M, De Vuyst L, Weckx S (2019) Shotgun metagenomics of a water kefir fermentation ecosystem reveals a novel oenococcus species. Front Microbiol 10: 479. https://doi.org/10.3389/fmicb.2019.00479
|
| [42] |
Ziarno M, Bryś J, Kowalska E, et al. (2023) Effect of metabolic activity of lactic acid bacteria and propionibacteria on cheese protein digestibility and fatty acid profile. Sci Rep 13: 15363. https://doi.org/10.1038/s41598-023-42633-w
|
| [43] |
Karimkhani MM, Jamshidi A, Nasrollahzadeh M, et al. (2024) Fermentation of Rubus dolichocarpus juice using Lactobacillus gasseri and Lacticaseibacillus casei and protecting phenolic compounds by Stevia extract during cold storage. Sci Rep 14: 5711. https://doi.org/10.1038/s41598-024-56235-7
|
| [44] |
Khan J, Sheoran S, Khan W, et al. (2020) Metabolic differentiation and quantification of gymnemic acid in GYMNEMA SYLVESTRE (Retz.) R.Br. ex Sm. leaf extract and its fermented products. Phytochem Anal 31: 488-500. https://doi.org/10.1002/pca.2912
|
| [45] |
Sagdic O, Ozturk I, Yapar N, et al. (2014) Diversity and probiotic potentials of lactic acid bacteria isolated from gilaburu, a traditional Turkish fermented European cranberrybush (Viburnum opulus L.) fruit drink. Food Res Int 64: 537-545. https://doi.org/10.1016/j.foodres.2014.07.045
|
| [46] |
Küçükgöz K, Kruk M, Kołożyn-Krajewska D, et al. (2024) Investigating the probiotic potential of vegan puree mixture: viability during simulated digestion and bioactive compound bioaccessibility. Nutrients 16: 561. https://doi.org/10.3390/nu16040561
|
| [47] |
Tsai-Hsin C, Shwu-Jene T, Tsung-Yen W, et al. (2013) Improvement in antioxidant activity, angiotensin-converting enzyme inhibitory activity and in vitro cellular properties of fermented pepino milk by Lactobacillus strains containing the glutamate decarboxylase gene. J Sci Food Agric 93: 859-866. https://doi.org/10.1002/jsfa.5809
|
| [48] |
Roman P, Carrillo-Trabalón F, Sánchez-Labraca N, et al. (2018) Are probiotic treatments useful on fibromyalgia syndrome or chronic fatigue syndrome patients? A systematic review. Benef Microbes 9: 603-611. https://doi.org/10.3920/bm2017.0125
|
| [49] |
Lee SH, Cho DY, Lee SH, et al. (2019) A randomized clinical trial of synbiotics in irritable bowel syndrome: dose-dependent effects on gastrointestinal symptoms and fatigue. Korean J Fam Med 40: 2. https://doi.org/10.4082/kjfm.17.0064
|
| [50] |
Peng M, Tabashsum Z, Patel P, et al. (2020) Prevention of enteric bacterial infections and modulation of gut microbiota with conjugated linoleic acids producing Lactobacillus in mice. Gut Microbes 11: 433-452. https://doi.org/10.1080/19490976.2019.1638724
|
| [51] |
Kumar MR, Yeap SK, Mohamad NE, et al. (2021) Metagenomic and phytochemical analyses of kefir water and its subchronic toxicity study in BALB/c mice. BMC Complement Med Ther 21: 183. https://doi.org/10.1186/s12906-021-03358-3
|
| [52] |
García-Ruiz A, Bartolomé B, Cueva C, et al. (2009) Inactivation of oenological lactic acid bacteria (Lactobacillus hilgardii and Pediococcus pentosaceus) by wine phenolic compounds. J Appl Microbiol 107: 1042-1053. https://doi.org/10.1111/j.1365-2672.2009.04287.x
|
| [53] |
Jurášková D, Ribeiro SC, Silva CCG (2022) Exopolysaccharides produced by lactic acid bacteria: from biosynthesis to health-promoting properties. Foods 11: 156. https://doi.org/10.3390/foods11020156
|
| [54] |
Wang Y, Wu Y, Sailike J, et al. (2020) Fourteen composite probiotics alleviate type 2 diabetes through modulating gut microbiota and modifying M1/M2 phenotype macrophage in db/db mice. Pharmacol Res 161: 105150. https://doi.org/10.1016/j.phrs.2020.105150
|
| [55] |
Yamaguchi T, Yasui K, Fujii S, et al. (2023) Lentilactobacillus hilgardii H-50 strongly inhibits lipopolysaccharide-induced inflammatory responses in mouse splenocytes via its specific surface layer proteins. J Appl Microbiol 134: lxad021. https://doi.org/10.1093/jambio/lxad021
|
| [56] |
Xia T, Wang T, Sun J, et al. (2022) Modulation of fermentation quality and metabolome in co-ensiling of sesbania cannabina and sweet sorghum by lactic acid bacterial inoculants. Front Microbiol 13: 851271. https://doi.org/10.3389/fmicb.2022.851271
|
| [57] |
Campos FM, Couto JA, Figueiredo AR, et al. (2009) Cell membrane damage induced by phenolic acids on wine lactic acid bacteria. Int J Food Microbiol 135: 144-151. https://doi.org/10.1016/j.ijfoodmicro.2009.07.031
|
| [58] |
Campos FM, Figueiredo AR, Hogg TA, et al. (2009) Effect of phenolic acids on glucose and organic acid metabolism by lactic acid bacteria from wine. Food Microbiol 26: 409-414. https://doi.org/10.1016/j.fm.2009.01.006
|
| [59] |
Xu D, Bechtner J, Behr J, et al. (2019) Lifestyle of Lactobacillus hordei isolated from water kefir based on genomic, proteomic and physiological characterization. Int J Food Microbiol 290: 141-149. https://doi.org/10.1016/j.ijfoodmicro.2018.10.004
|
| [60] |
Bechtner J, Xu D, Behr J, et al. (2019) Proteomic analysis of Lactobacillus nagelii in the presence of Saccharomyces cerevisiae isolated from water kefir and comparison with Lactobacillus hordei. Front Microbiol 10: 325. https://doi.org/10.3389/fmicb.2019.00325
|
| [61] |
Coniglio S, Shumskaya M, Vassiliou E (2023) Unsaturated fatty acids and their immunomodulatory properties. Biology 12: 279. https://doi.org/10.3390/biology12020279
|
| [62] |
Carasi P, Racedo SM, Jacquot C, et al. (2015) Impact of Kefir Derived Lactobacillus kefiri on the mucosal immune response and gut microbiota. J Immunol Res 2015: 1-12. https://doi.org/10.1155/2015/361604
|
| [63] |
Vieira LV, De Sousa LM, Maia TAC, et al. (2021) Milk Kefir therapy reduces inflammation and alveolar bone loss on periodontitis in rats. Biomed Pharmacother 139: 111677. https://doi.org/10.1016/j.biopha.2021.111677
|
| [64] |
Toscano M, De Grandi R, Miniello VL, et al. (2017) Ability of Lactobacillus kefiri LKF01 (DSM32079) to colonize the intestinal environment and modify the gut microbiota composition of healthy individuals. Dig Liver Dis 49: 261-267. https://doi.org/10.1016/j.dld.2016.11.011
|
| [65] |
Riaz Rajoka MS, Mehwish HM, Fang H, et al. (2019) Characterization and anti-tumor activity of exopolysaccharide produced by Lactobacillus kefiri isolated from Chinese kefir grains. J Funct Foods 63: 103588. https://doi.org/10.1016/j.jff.2019.103588
|
| [66] |
Yerlikaya O, Akan E, Kinik Ö (2022) The metagenomic composition of water kefir microbiota. Int J Gastron Food Sci 30: 100621. https://doi.org/10.1016/j.ijgfs.2022.100621
|
| [67] | Cunha C, Uecker JN, Jaskulski IB, et al. Probiotic characterization and safety assessment of Lactococcus Lactis Subsp. Lactis R7 isolated from ricotta cheese, in review. (2021). https://doi.org/10.21203/rs.3.rs-1135986/v1 |
| [68] |
Kleerebezem M, Bachmann H, van Pelt-KleinJan E, et al. (2020) Lifestyle, metabolism and environmental adaptation in Lactococcus lactis. FEMS Microbiol Rev 44: 804-820. https://doi.org/10.1093/femsre/fuaa033
|
| [69] |
Ayyash M, Olaimat A, Al-Nabulsi A, et al. (2020) Bioactive properties of novel probiotic Lactococcus lactis fermented camel sausages: cytotoxicity, angiotensin converting enzyme inhibition, antioxidant capacity, and antidiabetic activity. Food Sci Anim Resour 40: 155-171. https://doi.org/10.5851/kosfa.2020.e1
|
| [70] |
Nishitani Y, Tanoue T, Yamada K, et al. (2009) Lactococcus lactis subsp. cremoris FC alleviates symptoms of colitis induced by dextran sulfate sodium in mice. Int Immunopharmacol 9: 1444-1451. https://doi.org/10.1016/j.intimp.2009.08.018
|
| [71] |
Luerce TD, Gomes-Santos AC, Rocha CS, et al. (2014) Anti-inflammatory effects of Lactococcus lactis NCDO 2118 during the remission period of chemically induced colitis. Gut Pathog 6: 33. https://doi.org/10.1186/1757-4749-6-33
|
| [72] |
Li P, Xu Y, Cao Y, et al. (2022) Polypeptides Isolated from Lactococcus lactis Alleviates lipopolysaccharide (lps)-induced inflammation in Ctenopharyngodon idella. Int J Mol Sci 23: 6733. https://doi.org/10.3390/ijms23126733
|
| [73] |
Huang C, Kok J (2020) Editing of the proteolytic system of Lactococcus lactis increases its bioactive potential. Appl Environ Microbiol 86: e01319-20. https://doi.org/10.1128/AEM.01319-20
|
| [74] |
Egea MB, Santos DCD, Oliveira Filho JGD, et al. (2022) A review of nondairy kefir products: their characteristics and potential human health benefits. Crit Rev Food Sci Nutr 62: 1536-1552. https://doi.org/10.1080/10408398.2020.1844140
|
| [75] |
Kim S, Kim Y, Lee S, et al. (2022) Live biotherapeutic Lactococcus lactis GEN3013 enhances antitumor efficacy of cancer treatment via modulation of cancer progression and immune system. Cancers 14: 4083. https://doi.org/10.3390/cancers14174083
|
| [76] |
De Castro CP, Drumond MM, Batista VL, et al. (2018) Vector development timeline for mucosal vaccination and treatment of disease using Lactococcus lactis and design approaches of next generation food grade plasmids. Front Microbiol 9: 1805. https://doi.org/10.3389/fmicb.2018.01805
|
| [77] |
Benbouziane B, Ribelles P, Aubry C, et al. (2013) Development of a Stress-Inducible Controlled Expression (SICE) system in Lactococcus lactis for the production and delivery of therapeutic molecules at mucosal surfaces. J Biotechnol 168: 120-129. https://doi.org/10.1016/j.jbiotec.2013.04.019
|
| [78] |
Martín R, Martín R, Chain F, et al. (2014) Effects in the use of a genetically engineered strain of Lactococcus lactis delivering in situ IL-10 as a therapy to treat low-grade colon inflammation. Hum Vaccines Immunother 10: 1611-1621. https://doi.org/10.4161/hv.28549
|
| [79] |
Frossard CP, Steidler L, Eigenmann PA (2007) Oral administration of an IL-10–secreting Lactococcus lactis strain prevents food-induced IgE sensitization. J Allergy Clin Immunol 119: 952-959. https://doi.org/10.1016/j.jaci.2006.12.615
|
| [80] |
Zurita-Turk M, Mendes Souza B, Prósperi De Castro C, et al. (2020) Attenuation of intestinal inflammation in IL-10 deficient mice by a plasmid carrying Lactococcus lactis strain. BMC Biotechnol 20: 38. https://doi.org/10.1186/s12896-020-00631-0
|
| [81] |
Zurita-Turk M, Del Carmen S, Santos AC, et al. (2014) Lactococcus lactiscarrying the pValac DNA expression vector coding for IL-10 reduces inflammation in a murine model of experimental colitis. BMC Biotechnol 14: 73. https://doi.org/10.1186/1472-6750-14-73
|
| [82] |
Wang J, Tian M, Li W, et al. (2019) Preventative delivery of IL-35 by Lactococcus lactis ameliorates DSS-induced colitis in mice. Appl Microbiol Biotechnol 103: 7931-7941. https://doi.org/10.1007/s00253-019-10094-9
|
| [83] |
Bermúdez-Humarán LG, Langella P, Cortes-Perez NG, et al. (2003) Intranasal immunization with recombinant Lactococcus lactis secreting murine interleukin-12 enhances antigen-specific Th1 cytokine production. Infect Immun 71: 1887-1896. https://doi.org/10.1128/IAI.71.4.1887-1896.2003
|
| [84] |
Koh WY, Uthumporn U, Rosma A, et al. (2018) Fermented pumpkin-based beverage inhibits key enzymes of carbohydrate digesting and extenuates postprandial hyperglycemia in type-2 diabetic rats. Acta Aliment 47: 495-503. https://doi.org/10.1556/066.2018.47.4.13
|
| [85] |
Chen Y, Lin Y, Lin J, et al. (2018) Sugary kefir strain Lactobacillus mali APS1 ameliorated hepatic steatosis by regulation of SIRT-1/Nrf-2 and gut microbiota in rats. Mol Nutr Food Res 62: 1700903. https://doi.org/10.1002/mnfr.201700903
|
| [86] |
Lin YC, Chen YT, Hsieh HH, et al. (2016) Effect of Lactobacillus mali APS1 and L. kefiranofaciens M1 on obesity and glucose homeostasis in diet-induced obese mice. J Funct Foods 23: 580-589. https://doi.org/10.1016/j.jff.2016.03.015
|
| [87] |
Neville BA, Forde BM, Claesson MJ, et al. (2012) Characterization of pro-inflammatory flagellin proteins produced by Lactobacillus ruminis and related motile lactobacilli. PLoS ONE 7: e40592. https://doi.org/10.1371/journal.pone.0040592
|
| [88] |
Lin YC, Chen YT, Li KY, et al. (2020) Investigating the mechanistic differences of obesity-inducing Lactobacillus kefiranofaciens M1 and Anti-obesity Lactobacillus mali APS1 by microbolomics and metabolomics. Front Microbiol 11: 1454. https://doi.org/10.3389/fmicb.2020.01454
|
| [89] |
Hooi SL, Dwiyanto J, Toh KY, et al. (2023) The microbial composition and functional roles of different kombucha products in Singapore. CyTA-J Food 21: 269-274. https://doi.org/10.1080/19476337.2023.2190794
|
| [90] |
Dong K, Li W, Xu Q, et al. (2023) Exploring the correlation of metabolites changes and microbial succession in solid-state fermentation of Sichuan Sun-dried vinegar. BMC Microbiol 23: 197. https://doi.org/10.1186/s12866-023-02947-1
|
| [91] |
Rivera A, Becerra-Martinez E, Pacheco-Hernández Y, et al. (2020) Synergistic hypolipidemic and hypoglycemic effects of mixtures of Lactobacillus nagelii/betanin in a mouse model. Trop J Pharm Res 19: 1269-1276. https://doi.org/10.4314/tjpr.v19i6.23
|
| [92] |
Yang J, Lagishetty V, Kurnia P, et al. (2022) Microbial and chemical profiles of commercial kombucha products. Nutrients 14: 670. https://doi.org/10.3390/nu14030670
|
| [93] |
Mantzourani I, Terpou A, Bekatorou A, et al. (2022) Valorization of lactic acid fermentation of pomegranate juice by an acid tolerant and potentially probiotic lab isolated from kefir grains. Fermentation 8: 142. https://doi.org/10.3390/fermentation8040142
|
| [94] |
Li Y, Song H, Zhang Z, et al. (2024) Effects of fermentation with different probiotics on the quality, isoflavone content, and flavor of okara beverages. Food Sci Nutr 12: 2619-2633. https://doi.org/10.1002/fsn3.3944
|
| [95] |
Kim YB, Park J, Lee HG, et al. (2024) Dietary probiotic Lacticaseibacillus paracasei NSMJ56 modulates gut immunity and microbiota in laying hens. Poult Sci 103: 103505. https://doi.org/10.1016/j.psj.2024.103505
|
| [96] |
Lee W, Im H, Lee YB, et al. (2024) Protective effect of soy germ-fermented postbiotics derived from Lacticaseibacillus paracasei DCF0429 (SGPB-DCF0429) in human reconstituted skin. J Funct Foods 113: 106023. https://doi.org/10.1016/j.jff.2024.106023
|
| [97] |
Bhat B, Bajaj BK (2019) Hypocholesterolemic potential and bioactivity spectrum of an exopolysaccharide from a probiotic isolate Lactobacillus paracasei M7. Bioact Carbohydr Diet Fibre 19: 100191. https://doi.org/10.1016/j.bcdf.2019.100191
|
| [98] |
Balzaretti S, Taverniti V, Guglielmetti S, et al. (2017) A novel rhamnose-rich hetero-exopolysaccharide isolated from Lactobacillus paracasei DG activates THP-1 human monocytic cells. Appl Environ Microbiol 83: e02702-16. https://doi.org/10.1128/AEM.02702-16
|
| [99] |
Ahn H, Lee G, Lee W, et al. (2023) Evaluation of probiotic and anti-inflammatory properties of bacteriocinogenic Pediococcus acidilactici HW01 and Leuconostoc citreum HW02 from malted barley. Chem Biol Technol Agric 10: 49. https://doi.org/10.1186/s40538-023-00425-4
|
| [100] |
Han HS, Soundharrajan I, Valan Arasu M, et al. (2023) Leuconostoc citreum inhibits adipogenesis and lipogenesis by inhibiting p38 MAPK/Erk 44/42 and stimulating AMPKα signaling pathways. Int J Mol Sci 24: 7367. https://doi.org/10.3390/ijms24087367
|
| [101] |
Muthusamy K, Han HS, Soundharrajan I, et al. (2023) A novel strain of probiotic Leuconostoc citreum inhibits infection-causing bacterial pathogens. Microorganisms 11: 469. https://doi.org/10.3390/microorganisms11020469
|
| [102] |
Son J, Jeong KJ (2022) Engineering of Leuconostoc citreum for efficient bioconversion of soy isoflavone glycosides to their aglycone forms. Int J Mol Sci 23: 9568. https://doi.org/10.3390/ijms23179568
|
| [103] |
Li Y, Xiao L, Tian J, et al. (2022) Structural characterization, rheological properties and protection of oxidative damage of an exopolysaccharide from Leuconostoc citreum 1.2461 fermented in soybean whey. Foods 11: 2283. https://doi.org/10.3390/foods11152283
|
| [104] |
Wang Y, Du R, Qiao X, et al. (2020) Optimization and characterization of exopolysaccharides with a highly branched structure extracted from Leuconostoc citreum B-2. Int J Biol Macromol 142: 73-84. https://doi.org/10.1016/j.ijbiomac.2019.09.071
|
| [105] |
Kim M, Jang JK, Park YS (2021) Production optimization, structural analysis, and prebiotic- and anti-inflammatory effects of gluco-oligosaccharides produced by Leuconostoc lactis SBC001. Microorganisms 9: 200. https://doi.org/10.3390/microorganisms9010200
|
| [106] |
Chang-Liao WP, Lee A, Chiu YH, et al. (2020) Isolation of a Leuconostoc mesenteroides strain with anti-porcine epidemic diarrhea virus activities from kefir grains. Front Microbiol 11: 1578. https://doi.org/10.3389/fmicb.2020.01578
|
| [107] |
Luan C, Yan J, Jiang N, et al. (2022) Leuconostoc mesenteroides LVBH107 antibacterial activity against porphyromonas gingivalis and anti-inflammatory activity against P. gingivalis lipopolysaccharide-stimulated RAW 264.7 cells. Nutrients 14: 2584. https://doi.org/10.3390/nu14132584
|
| [108] |
Moon HJ, Oh SH, Park KB, et al. (2023) Kimchi and Leuconostoc mesenteroides DRC 1506 alleviate dextran sulfate sodium (dss)-induced colitis via attenuating inflammatory responses. Foods 12: 584. https://doi.org/10.3390/foods12030584
|
| [109] |
Zhang Q, Wang J, Sun Q, et al. (2021) Characterization and antioxidant activity of released exopolysaccharide from potential probiotic Leuconostoc mesenteroides LM187. J Microbiol Biotechnol 31: 1144-1153. https://doi.org/10.4014/jmb.2103.03055
|
| [110] |
Li Y, Liu Y, Cao C, et al. (2020) Extraction and biological activity of exopolysaccharide produced by Leuconostoc mesenteroides SN-8. Int J Biol Macromol 157: 36-44. https://doi.org/10.1016/j.ijbiomac.2020.04.150
|
| [111] |
Taylan O, Yilmaz MT, Dertli E (2019) Partial characterization of a levan type exopolysaccharide (EPS) produced by Leuconostoc mesenteroides showing immunostimulatory and antioxidant activities. Int J Biol Macromol 136: 436-444. https://doi.org/10.1016/j.ijbiomac.2019.06.078
|
| [112] |
Hwang JE, Kim KT, Paik HD (2019) Improved antioxidant, anti-inflammatory, and anti-adipogenic properties of hydroponic ginseng fermented by Leuconostoc mesenteroides KCCM 12010P. Molecules 24: 3359. https://doi.org/10.3390/molecules24183359
|
| [113] |
Daliri EB, Choi S, Cho B, et al. (2019) Biological activities of a garlic–Cirsium setidens Nakai blend fermented with Leuconostoc mesenteroides. Food Sci Nutr 7: 2024-2032. https://doi.org/10.1002/fsn3.1032
|
| [114] |
Li Y, Liu L, Yu X, et al. (2019) Transglycosylation improved caffeic acid phenethyl ester anti-inflammatory activity and water solubility by Leuconostoc mesenteroides dextransucrase. J Agric Food Chem 67: 4505-4512. https://doi.org/10.1021/acs.jafc.9b01143
|
| [115] | Murtaza G, Karim S, Akram MR, et al. (2014) Caffeic acid phenethyl ester and therapeutic potentials. BioMed Res Int 2014: 1-9. https://doi.org/10.1155/2014/145342 |
| [116] |
Woo HJ, Kang HK, Nguyen TTH, et al. (2012) Synthesis and characterization of ampelopsin glucosides using dextransucrase from Leuconostoc mesenteroides B-1299CB4: Glucosylation enhancing physicochemical properties. Enzyme Microb Technol 51: 311-318. https://doi.org/10.1016/j.enzmictec.2012.07.014
|
| [117] |
Ruiz-de-Villa C, Poblet M, Bordons A, et al. (2023) Comparative study of inoculation strategies of Torulaspora delbrueckii and Saccharomyces cerevisiae on the performance of alcoholic and malolactic fermentations in an optimized synthetic grape must. Int J Food Microbiol 404: 110367. https://doi.org/10.1016/j.ijfoodmicro.2023.110367
|
| [118] |
Su J, Wang T, Li YY, et al. (2015) Antioxidant properties of wine lactic acid bacteria: Oenococcus oeni. Appl Microbiol Biotechnol 99: 5189-5202. https://doi.org/10.1007/s00253-015-6425-4
|
| [119] |
Luciana Del Valle R, Carmen M, María José R-V, et al. (2022) Utilization of Oenococcus oeni strains to ferment grape juice: Metabolic activities and beneficial health potential. Food Microbiol 101: 103895. https://doi.org/10.1016/j.fm.2021.103895
|
| [120] |
Kristof I, Ledesma SC, Apud GR, et al. (2023) Oenococcus oeni allows the increase of antihypertensive and antioxidant activities in apple cider. Heliyon 9: e16806. https://doi.org/10.1016/j.heliyon.2023.e16806
|
| [121] |
Foligné B, Dewulf J, Breton J, et al. (2010) Probiotic properties of non-conventional lactic acid bacteria: Immunomodulation by Oenococcus oeni. Int J Food Microbiol 140: 136-145. https://doi.org/10.1016/j.ijfoodmicro.2010.04.007
|
| [122] |
Hossain S, Khetra Y, Dularia C, et al. (2023) Symbiotic fermentation study of Acetobacter orientalis and lactic acid bacteria for lactobionic acid enriched yoghurt production. Food Biosci 53: 102612. https://doi.org/10.1016/j.fbio.2023.102612
|
| [123] |
Gamba RR, Yamamoto S, Abdel-Hamid M, et al. (2020) Chemical, microbiological, and functional characterization of kefir produced from cow's milk and soy milk. Int J Microbiol 2020: 1-11. https://doi.org/10.1155/2020/7019286
|
| [124] |
Aligita W, Singgih M, Sutrisno E, et al. (2023) Hepatoprotective properties of water kefir: A traditional fermented drink and its potential role. Int J Prev Med 14: 93. https://doi.org/10.4103%2Fijpvm.ijpvm_29_22
|
| [125] |
Cai C, Li Z, Lu J, et al. (2023) Effects of acetoin on growth performance, digestive function, antioxidant status, and immune capacity of largemouth bass (Micropterus salmoides). Aquac Res 2023: 1-12. https://doi.org/10.1155/2023/6114525
|
| [126] |
Michiels J, Truffin D, Majdeddin M, et al. (2023) Gluconic acid improves performance of newly weaned piglets associated with alterations in gut microbiome and fermentation. Porc Health Manag 9: 10. https://doi.org/10.1186/s40813-023-00305-1
|
| [127] |
Fischer F, Romero R, Hellhund A, et al. (2020) Dietary cellulose induces anti-inflammatory immunity and transcriptional programs via maturation of the intestinal microbiota. Gut Microbes 12: 1829962. https://doi.org/10.1080/19490976.2020.1829962
|
| [128] |
White KM, Matthews MK, Hughes RC, et al. (2018) A metagenome-wide association study and arrayed mutant library confirm Acetobacter lipopolysaccharide genes are necessary for association with Drosophila melanogaster. G3 GenesGenomesGenetics 8: 1119-1127. https://doi.org/10.1534/g3.117.300530
|
| [129] |
Kong Y, Wang L, Jiang B (2021) The role of gut microbiota in aging and aging related neurodegenerative disorders: insights from drosophila model. Life 11: 855. https://doi.org/10.3390/life11080855
|
| [130] |
Batista LL, Malta SM, Guerra Silva HC, et al. (2021) Kefir metabolites in a fly model for Alzheimer's disease. Sci Rep 11: 11262. https://doi.org/10.1038/s41598-021-90749-8
|
| [131] |
Kim D, Kim H, Seo K (2020) Microbial composition of Korean kefir and antimicrobial activity of Acetobacter fabarum DH1801. J Food Saf 40: e12728. https://doi.org/10.1111/jfs.12728
|
| [132] |
Martínez-Torres A, Gutiérrez-Ambrocio S, Heredia-del-Orbe P, et al. (2017) Inferring the role of microorganisms in waterkefirfermentations. Int J Food Sci Technol 52: 559-571. https://doi.org/10.1111/ijfs.13312
|
| [133] |
Uchida K, Akashi K, Motoshima H, et al. (2009) Microbiota analysis of Caspian Sea yogurt, a ropy fermented milk circulated in Japan. Anim Sci J 80: 187-192. https://doi.org/10.1111/j.1740-0929.2008.00607.x
|
| [134] |
Eble H, Joswig M, Lamberti L, et al. (2023) Master regulators of biological systems in higher dimensions. Proc Natl Acad Sci 120: e2300634120. https://doi.org/10.1073/pnas.2300634120
|
| [135] |
Hou Z, Sun L, Wang D, et al. (2020) Production of 2-keto-gluconic acid from glucose by immobilized Pseudomonas plecoglossicida resting cells. 3 Biotech 10: 253. https://doi.org/10.1007/s13205-020-02243-z
|
| [136] |
Goderska K (2019) The antioxidant and prebiotic properties of lactobionic acid. Appl Microbiol Biotechnol 103: 3737-3751. https://doi.org/10.1007/s00253-019-09754-7
|
| [137] |
Arrieta-Echeverri MC, Fernandez GJ, Duarte-Riveros A, et al. (2023) Multi-omics characterization of the microbial populations and chemical space composition of a water kefir fermentation. Front Mol Biosci 10: 1223863. https://doi.org/10.3389/fmolb.2023.1223863
|
| [138] | Kusnadi J, Tirtania AR, Arumingtyas EL (2023) Antioxidant activity, physicochemical characterisation and antibacterial properties of caspian sea yoghurt enriched with ginger and sappanwood extracts. Trop J Nat Prod Res 7. http://www.doi.org/10.26538/tjnpr/v7i3.11 |
| [139] |
Lee S, Lee JA, Park GG, et al. (2017) Semi-continuous fermentation of onion vinegar and its functional properties. Molecules 22: 1313. https://doi.org/10.3390/molecules22081313
|
| [140] |
Lim JM, Lee SH, Jeong DY, et al. (2022) Significance of LED lights in enhancing the production of vinegar using Acetobacter pasteurianus AP01. Prep Biochem Biotechnol 52: 38-47. https://doi.org/10.1080/10826068.2021.1907406
|
| [141] |
Taweekasemsombut S, Tinoi J, Mungkornasawakul P, et al. (2021) Thai rice vinegars: production and biological properties. Appl Sci 11: 5929. https://doi.org/10.3390/app11135929
|
| [142] |
Liu X, Zhang L, Cao C, et al. (2023) Biorefining process of agricultural onions to functional vinegar. Prep Biochem Biotechnol 53: 424-432. https://doi.org/10.1080/10826068.2022.2098321
|
| [143] |
Luang-In V, Saengha W, Yotchaisarn M, et al. (2018) Microbial strains and bioactive exopolysaccharide producers from Thai Water Kefir. Microbiol Biotechnol Lett 46: 403-415. https://doi.org/10.4014/mbl.1804.04019
|
| [144] |
Wen X, Wang Z, Liu Q, et al. (2023) Acetobacter pasteurianus BP2201 alleviates alcohol-induced hepatic and neuro-toxicity and modulate gut microbiota in mice. Microb Biotechnol 16: 1834-1857. https://doi.org/10.1111/1751-7915.14303
|
| [145] |
Ankrah NYD, Barker BE, Song J, et al. (2021) Predicted metabolic function of the gut microbiota of Drosophila melanogaster. mSystems 6: e01369-20. https://doi.org/10.1128/mSystems.01369-20
|
| [146] |
Huang JH, Douglas AE (2015) Consumption of dietary sugar by gut bacteria determines Drosophila lipid content. Biol Lett 11: 20150469. https://doi.org/10.1098/rsbl.2015.0469
|
| [147] | Tokatli Demi̇Rok N, Alpaslan M, Yikmiş S (2023) Some lactobacillus, leuconostoc and acetobacter strains in traditional turkish yoghurt, cheese, kefir samples as a probiotic candidate. Int J Agric Environ Food Sci 7: 326-334. https://doi.org/10.31015/jaefs.2023.2.10 |
| [148] |
Purutoğlu K, İspirli H, Yüzer MO, et al. (2020) Diversity and functional characteristics of lactic acid bacteria from traditional kefir grains. Int J Dairy Technol 73: 57-66. https://doi.org/10.1111/1471-0307.12633
|
| [149] |
Milani C, Lugli GA, Duranti S, et al. (2014) Genomic encyclopedia of type strains of the genus bifidobacterium. Appl Environ Microbiol 80: 6290-6302. https://doi.org/10.1128/AEM.02308-14
|
| [150] |
Laureys D, Cnockaert M, De Vuyst L, et al. (2016) Bifidobacterium aquikefiri sp. nov., isolated from water kefir. Int J Syst Evol Microbiol 66: 1281-1286. https://doi.org/10.1099/ijsem.0.000877
|
| [151] |
Laureys D, Van Jean A, Dumont J, et al. (2017) Investigation of the instability and low water kefir grain growth during an industrial water kefir fermentation process. Appl Microbiol Biotechnol 101: 2811-2819. https://doi.org/10.1007/s00253-016-8084-5
|
| [152] |
Patel SH, Tan JP, Börner RA, et al. (2022) A temporal view of the water kefir microbiota and flavour attributes. Innov Food Sci Emerg Technol 80: 103084. https://doi.org/10.1016/j.ifset.2022.103084
|
| [153] |
Manosalva C, Quiroga J, Hidalgo AI, et al. (2022) Role of lactate in inflammatory processes: friend or foe. Front Immunol 12: 808799. https://doi.org/10.3389/fimmu.2021.808799
|
| [154] |
Cavone L, Calosi L, Cinci L, et al. (2012) Topical mannitol reduces inflammatory edema in a rat model of arthritis. Pharmacology 89: 18-21. https://doi.org/10.1159/000335094
|
| [155] |
Pelle E, Mammone T, Marenus K, et al. (2003) Ultraviolet-b-induced oxidative DNA base damage in primary normal human epidermal keratinocytes and inhibition by a hydroxyl radical scavenger. J Invest Dermatol 121: 177-183. https://doi.org/10.1046/j.1523-1747.2003.12330.x
|
| [156] |
Ratter JM, Rooijackers HMM, Hooiveld GJ, et al. (2018) In vitro and in vivo effects of lactate on metabolism and cytokine production of human primary pbmcs and monocytes. Front Immunol 9: 2564. https://doi.org/10.3389/fimmu.2018.02564
|
| [157] |
Eckel VPL, Vogel RF (2020) Genomic and physiological insights into the lifestyle of Bifidobacterium species from water kefir. Arch Microbiol 202: 1627-1637. https://doi.org/10.1007/s00203-020-01870-7
|
| [158] |
Grosser N, Oberle S, Berndt G, et al. (2004) Antioxidant action of l-alanine: heme oxygenase-1 and ferritin as possible mediators. Biochem Biophys Res Commun 314: 351-355. https://doi.org/10.1016/j.bbrc.2003.12.089
|
| [159] |
Pires RS, Braga PGS, Santos JMB, et al. (2021) l-Glutamine supplementation enhances glutathione peroxidase and paraoxonase-1 activities in HDL of exercising older individuals. Exp Gerontol 156: 111584. https://doi.org/10.1016/j.exger.2021.111584
|
| [160] | Moradi M, Moradi B, Hashemian AH, et al. (2022) Beneficial effect of L-Proline supplementation on the quality of human spermatozoa. Andrologia 54. https://doi.org/10.1111/and.14486 |
| [161] |
Abbasi M, Taheri Mirghaed A, Hoseini SM, et al. (2023) Effects of dietary glycine supplementation on growth performance, immunological, and erythrocyte antioxidant parameters in common carp, Cyprinus carpio. Animals 13: 412. https://doi.org/10.3390/ani13030412
|
| [162] |
Zhang D, Nie S, Xie M, et al. (2020) Antioxidant and antibacterial capabilities of phenolic compounds and organic acids from Camellia oleifera cake. Food Sci Biotechnol 29: 17-25.
|
| [163] |
Schöpping M, Zeidan AA, Franzén CJ (2022) Stress response in bifidobacteria. Microbiol Mol Biol Rev 86: e00170-21. https://doi.org/10.1007%2Fs10068-019-00637-1
|
| [164] | Younus H (2018) Therapeutic potentials of superoxide dismutase. Int J Health Sci 12: 88. Available from: http://www.ncbi.nlm.nih.gov/pmc/articles/pmc5969776/ |
| [165] | Sarıkaya E, Doğan S (2020) Glutathione peroxidase in health and diseases. Glutathione System and Oxidative Stress in Health and Disease . IntechOpen. https://doi.org/10.5772/intechopen.91009 |
| [166] |
Gulitz A, Stadie J, Ehrmann MA, et al. (2013) Comparative phylobiomic analysis of the bacterial community of water kefir by 16S rRNA gene amplicon sequencing and ARDRA analysis. J Appl Microbiol 114: 1082-1091. https://doi.org/10.1111/jam.12124
|
| [167] |
Delcenserie V, Gavini F, Beerens H, et al. (2007) Description of a new species, Bifidobacterium crudilactis sp. nov., isolated from raw milk and raw milk cheeses. Syst Appl Microbiol 30: 381-389. https://doi.org/10.1016/j.syapm.2007.01.004
|
| [168] |
Delcenserie V, Taminiau B, Gavini F, et al. (2013) Detection and characterization of Bifidobacterium crudilactis and B. mongoliense able to grow during the manufacturing process of French raw milk cheeses. BMC Microbiol 13: 239. https://doi.org/10.1186/1471-2180-13-239
|
| [169] |
Bondue P, Lebrun S, Taminiau B, et al. (2020) Effect of Bifidobacterium crudilactis and 3′-sialyllactose on the toddler microbiota using the SHIME® model. Food Res Int 138: 109755. https://doi.org/10.1016/j.foodres.2020.109755
|
| [170] |
Yao Y, Cai X, Fei W, et al. (2022) The role of short-chain fatty acids in immunity, inflammation and metabolism. Crit Rev Food Sci Nutr 62: 1-12. https://doi.org/10.1080/10408398.2020.1854675
|
| [171] |
Wu J, Chen N, Grau E, et al. (2024) Short chain fatty acids inhibit corneal inflammatory responses to TLR ligands via the ocular G-protein coupled receptor 43. Ocul Surf 32: 48-57. https://doi.org/10.1016/j.jtos.2024.01.005
|
| [172] |
Jiang M, Li Z, Zhang F, et al. (2023) Butyrate inhibits iILC2-mediated lung inflammation via lung-gut axis in chronic obstructive pulmonary disease (COPD). BMC Pulm Med 23: 163. https://doi.org/10.1186/s12890-023-02438-z
|
| [173] |
Sato Y, Kanayama M, Nakajima S, et al. (2024) Sialyllactose enhances the short-chain fatty acid production and barrier function of gut epithelial cells via nonbifidogenic modification of the fecal microbiome in human adults. Microorganisms 12: 252. https://doi.org/10.3390/microorganisms12020252
|
| [174] |
Gökırmaklı Ç, Yüceer YK, Guzel-Seydim ZB (2023) Chemical, microbial, and volatile changes of water kefir during fermentation with economic substrates. Eur Food Res Technol 249: 1717-1728. https://doi.org/10.1007/s00217-023-04242-9
|
| [175] |
Laureys D, De Vuyst L (2014) Microbial species diversity, community dynamics, and metabolite kinetics of water kefir fermentation. Appl Environ Microbiol 80: 2564-2572. https://doi.org/10.1128/AEM.03978-13
|
| [176] | Raimondi S, Amaretti A, Leonardi A, et al. (2016) Conjugated linoleic acid production by bifidobacteria: screening, kinetic, and composition. BioMed Res Int 2016: 1-8. https://doi.org/10.1155/2016/8654317 |
| [177] |
Valenzuela CA, Baker EJ, Miles EA, et al. (2023) Conjugated linoleic acids have anti-inflammatory effects in cultured endothelial cells. Int J Mol Sci 24: 874. https://doi.org/10.3390/ijms24010874
|
| [178] |
Huang WC, Tu RS, Chen YL, et al. (2016) Conjugated linoleic acids suppress inflammatory response and ICAM-1 expression through inhibition of NF-κB and MAPK signaling in human bronchial epithelial cells. Food Funct 7: 2025-2033. https://doi.org/10.1039/C5FO01037C
|
| [179] |
Wijaya L, Sumerta IN, Napitupulu TP, et al. (2024) Cultural, nutritional and microbial perspectives of tuak, a traditional Balinese beverage. J Ethn Foods 11: 1-14. https://doi.org/10.1186/s42779-024-00221-x
|
| [180] | Rogers P, Jeon Y, Lee K, et al. (2007) Zymomonas mobilis for fuel ethanol and higher value products. Biofuels 263–288. https://doi.org/10.1007/10_2007_060 |
| [181] | Kim KH, Chung CB, Kim YH, et al. (2005) Cosmeceutical properties of levan produced by Zymomonas mobilis. J Cosmet Sci 56: 395-406. http://dx.doi.org/10.1111/j.1467-2494.2006.00314_2.x |
| [182] |
Dawes E, Ribbons D, Rees D (1966) Sucrose utilization by Zymomonas mobilis: formation of a levan. Biochem J 98: 804. https://doi.org/10.1042/bj0980804
|
| [183] | Scopes R (1987) Zymomonas as a source of diagnostic enzymes. Aust J Biotechnol 1: 58-63. Available from: https://scholar.google.com/scholar_lookup?&title=Zymomonas%20as%20a%20source%20of%20diagnostic%20enzymes&journal=Aust.%20J.%20Biotechnol&volume=1&issue=3&pages=58-63&publication_year=1987&author=Scopes%2CRK |
| [184] |
Uhlenbusch I, Sahm H, Sprenger GA (1991) Expression of an L-alanine dehydrogenase gene in Zymomonas mobilis and excretion of L-alanine. Appl Environ Microbiol 57: 1360-1366. https://doi.org/10.1128/aem.57.5.1360-1366.1991
|
| [185] | Kim JY, Shin SH, Chong HY, et al. Transformant for production of lactic acid of high optical purity and method for producing lactic acid using the same (2016). Available from: https://patents.google.com/patent/US9428775B2/en |
| [186] |
Misawa N, Yamano S, Ikenaga H (1991) Production of beta-carotene in Zymomonas mobilis and Agrobacterium tumefaciens by introduction of the biosynthesis genes from Erwinia uredovora. Appl Environ Microbiol 57: 1847-1849. https://doi.org/10.1128%2Faem.57.6.1847-1849.1991
|
| [187] |
Liebler DC, Stratton SP, Kaysen KL (1997) Antioxidant actions of β-carotene in liposomal and microsomal membranes: role of carotenoid-membrane incorporation and α-tocopherol. Arch Biochem Biophys 338: 244-250. https://doi.org/10.1006/abbi.1996.9822
|
| [188] | Yang Y, Li R, Hui J, et al. (2021) β-Carotene attenuates LPS-induced rat intestinal inflammation via modulating autophagy and regulating the JAK2/STAT3 and JNK/p38 MAPK signaling pathways. J Food Biochem 45. https://doi.org/10.1111/jfbc.13544 |
| [189] |
Tornabene TG, Holzer G, Bittner AS, et al. (1982) Characterization of the total extractable lipids of Zymomonas mobilis var. mobilis. Can J Microbiol 28: 1107-1118. https://doi.org/10.1139/m82-165
|
| [190] |
Carey VC, Ingram LO (1983) Lipid composition of Zymomonas mobilis: effects of ethanol and glucose. J Bacteriol 154: 1291-1300. https://doi.org/10.1128/jb.154.3.1291-1300.1983
|
| [191] |
Barrow KD, Collins JG, Rogers PL, et al. (1983) Lipid composition of an ethanol-tolerant strain of Zymomonas mobilis. Biochim Biophys Acta BBA-Lipids Lipid Metab 753: 324-330. https://doi.org/10.1016/0005-2760(83)90055-3
|
| [192] |
Khalil ASM, Giribabu N, Yelumalai S, et al. (2021) Myristic acid defends against testicular oxidative stress, inflammation, apoptosis: Restoration of spermatogenesis, steroidogenesis in diabetic rats. Life Sci 278: 119605. https://doi.org/10.1016/j.lfs.2021.119605
|
| [193] |
Zadeh Hashem E, Khodadadi M, Asadi F, et al. (2016) The antioxidant activity of palmitoleic acid on the oxidative stress parameters of palmitic acid in adult rat cardiomyocytes. Ann Mil Health Sci Res 14. https://doi.org/10.5812/amh.11467
|
| [194] |
Tsai YW, Lu CH, Chang RCA, et al. (2021) Palmitoleic acid ameliorates palmitic acid-induced proinflammation in J774A.1 macrophages via TLR4-dependent and TNF-α-independent signallings. Prostaglandins Leukot Essent Fatty Acids 169: 102270. https://doi.org/10.1016/j.plefa.2021.102270
|
| [195] |
Wang Z, Liang C, Li G, et al. (2007) Stearic acid protects primary cultured cortical neurons against oxidative stress. Acta Pharmacol Sin 28: 315-326. https://doi.org/10.1111/j.1745-7254.2007.00512.x
|
| [196] |
Pan PH, Lin SY, Ou YC, et al. (2010) Stearic acid attenuates cholestasis-induced liver injury. Biochem Biophys Res Commun 391: 1537-1542. https://doi.org/10.1016/j.bbrc.2009.12.119
|
| [197] |
Alves NFB, de Queiroz TM, de Almeida Travassos R, et al. (2017) Acute treatment with lauric acid reduces blood pressure and oxidative stress in spontaneously hypertensive rats. Basic Clin Pharmacol Toxicol 120: 348-353. https://doi.org/10.1111/bcpt.12700
|
| [198] | Dubo A, Dawud F, Umar I, et al. (2019) Lauric acid alleviates inflammation and structural changes in the lungs of type II diabetic male Wistar rats. J Afr Assoc Physiol Sci 7: 88-96. Available from: https://www.ajol.info/index.php/jaaps/article/view/192646 |
| [199] |
Tsoupras AB, Demopoulos CA, Pappas KM (2012) Platelet-activating factor detection, metabolism, and inhibitors in the ethanologenic bacterium Zymomonas mobilis. Eur J Lipid Sci Technol 114: 123-133. https://doi.org/10.1002/ejlt.201000489
|
| [200] |
Campos IA, Ximenes EA, Carvalho Júnior CHR, et al. (2013) Zymomonas mobilis culture protects against sepsis by modulating the inflammatory response, alleviating bacterial burden and suppressing splenocyte apoptosis. Eur J Pharm Sci 48: 1-8. https://doi.org/10.1016/j.ejps.2012.10.011
|
| [201] |
Diaz M, Kellingray L, Akinyemi N, et al. (2019) Comparison of the microbial composition of African fermented foods using amplicon sequencing. Sci Rep 9: 13863. https://doi.org/10.1038/s41598-019-50190-4
|
| [202] | Lindner P Atlas der mikroskopischen Grundlagen der Garungskunde, Tafel 68 (1928). Available from: https://scholar.google.com/scholar?hl=el&as_sdt=0%2C5&q=Lindner%2C+P.+%281928%29.+Atlas+der+mikroskopischen+Grundlagen+der+Garungskunde%2C+Tafel+68.&btnG= |
| [203] | Millis NF Some bacterial fermentations of cider (1951). Available from: https://scholar.google.com/scholar?hl=el&as_sdt=0%2C5&q=Millis%2C+N.+F.+%281951%29.+Some+bacterial+fermentations+of+cider.&btnG= |
| [204] |
Ruiz-Argueso T, Rodriguez-Navarro A (1975) Microbiology of ripening honey. Appl Microbiol 30: 893-896. https://doi.org/10.1128%2Fam.30.6.893-896.1975
|
| [205] |
Fentie EG, Jeong M, Emire SA, et al. (2022) Physicochemical properties, antioxidant activities and microbial communities of Ethiopian honey wine, Tej. Food Res Int 152: 110765. https://doi.org/10.1016/j.foodres.2021.110765
|
| [206] |
Fabricio MF, Mann MB, Kothe CI, et al. (2022) Effect of freeze-dried kombucha culture on microbial composition and assessment of metabolic dynamics during fermentation. Food Microbiol 101: 103889. https://doi.org/10.1016/j.fm.2021.103889
|
| [207] |
Kawamata Y, Toyotake Y, Ogiyama D, et al. (2021) Development of the original whey-based vinegar using rapeseed meal or wheat bran as a raw material for koji. J Food Process Preserv 45. https://doi.org/10.1111/jfpp.16097
|
| [208] |
Wang B, Rutherfurd-Markwick K, Liu N, et al. (2024) Evaluation of the probiotic potential of yeast isolated from kombucha in New Zealand. Curr Res Food Sci 8: 100711. https://doi.org/10.1016/j.crfs.2024.100711
|
| [209] | Cosmetic composition containing polyorganosiloxane-containing epsilon-polylysine polymer, and polyhydric alcohol, and production thereof. Available from: https://patents.google.com/patent/EP1604647A1/en |
| [210] |
Lam R, Lin ZX, Sviderskaya E, et al. (2014) Mechanistic studies of anti-hyperpigmentary compounds: elucidating their inhibitory and regulatory actions. Int J Mol Sci 15: 14649-14668. https://doi.org/10.3390/ijms150814649
|
| [211] |
Köhler S, Schmacht M, Troubounis AHL, et al. (2021) Tradition as a stepping stone for a microbial defined water kefir fermentation process: insights in cell growth, bioflavoring, and sensory perception. Front Microbiol 12: 732019. https://doi.org/10.3389/fmicb.2021.732019
|
| [212] |
Kim H, Hur S, Lim J, et al. (2023) Enhancement of the phenolic compounds and antioxidant activities of Kombucha prepared using specific bacterial and yeast. Food Biosci 56: 103431. https://doi.org/10.1016/j.fbio.2023.103431
|
| [213] |
Vigentini I, Romano A, Compagno C, et al. (2008) Physiological and oenological traits of different Dekkera/Brettanomyces bruxellensis strains under wine-model conditions. FEMS Yeast Res 8: 1087-1096. https://doi.org/10.1111/j.1567-1364.2008.00395.x
|
| [214] |
Silva LR, Andrade PB, Valentão P, et al. (2005) Analysis of non-coloured phenolics in red wine: Effect of Dekkera bruxellensis yeast. Food Chem 89: 185-189. https://doi.org/10.1016/j.foodchem.2004.02.019
|
| [215] |
Schifferdecker AJ, Dashko S, Ishchuk OP, et al. (2014) The wine and beer yeast Dekkera bruxellensis. Yeast 31: 323-332. https://doi.org/10.1002/yea.3023
|
| [216] |
Sun TY, Li JS, Chen C (2015) Effects of blending wheatgrass juice on enhancing phenolic compounds and antioxidant activities of traditional kombucha beverage. J Food Drug Anal 23: 709-718. https://doi.org/10.1016/j.jfda.2015.01.009
|
| [217] |
Lee J, Cho J, Kim J, et al. (2022) Evaluation of the fermentation characteristics and functionality of kombucha for commercialization. J Korean Soc Food Sci Nutr 51: 811-818. https://doi.org/10.3746/jkfn.2022.51.8.811
|
| [218] |
Tran T, Roullier-Gall C, Verdier F, et al. (2022) Microbial interactions in kombucha through the lens of metabolomics. Metabolites 12: 235. https://doi.org/10.3390/metabo12030235
|
| [219] |
Tran T, Billet K, Torres-Cobos B, et al. (2022) Use of a minimal microbial consortium to determine the origin of kombucha flavor. Front Microbiol 13: 836617. https://doi.org/10.3389/fmicb.2022.836617
|
| [220] |
Lalitha P, Parthiban A, Sachithanandam V, et al. (2021) Antibacterial and antioxidant potential of GC-MS analysis of crude ethyl acetate extract from the tropical mangrove plant Avicennia officinalis L. South Afr J Bot 142: 149-155. https://doi.org/10.1016/j.sajb.2021.06.023
|
| [221] |
Bellut K, Krogerus K, Arendt EK (2020) Lachancea fermentati strains isolated from kombucha: fundamental insights, and practical application in low alcohol beer brewing. Front Microbiol 11: 764. https://doi.org/10.3389/fmicb.2020.00764
|
| [222] |
Maciel NOP, Piló FB, Freitas LFD, et al. (2013) The diversity and antifungal susceptibility of the yeasts isolated from coconut water and reconstituted fruit juices in Brazil. Int J Food Microbiol 160: 201-205. https://doi.org/10.1016/j.ijfoodmicro.2012.10.012
|
| [223] |
Leuck AM, Rothenberger MK, Green JS (2014) Fungemia due to Lachancea fermentati: a case report. BMC Infect Dis 14: 250. https://doi.org/10.1186/1471-2334-14-250
|
| [224] |
Fakruddin Md, Hossain MdN, Ahmed MM (2017) Antimicrobial and antioxidant activities of Saccharomyces cerevisiae IFST062013, a potential probiotic. BMC Complement Altern Med 17: 64. https://doi.org/10.1186/s12906-017-1591-9
|
| [225] |
Eppinga H, Thio HB, Schreurs MWJ, et al. (2017) Depletion of Saccharomyces cerevisiae in psoriasis patients, restored by Dimethylfumarate therapy (DMF). PLOS ONE 12: e0176955. https://doi.org/10.1371/journal.pone.0176955
|
| [226] |
Gabrielli E, Pericolini E, Ballet N, et al. (2018) Saccharomyces cerevisiae-based probiotic as novel anti-fungal and anti-inflammatory agent for therapy of vaginal candidiasis. Benef Microbes 9: 219-230. https://doi.org/10.3920/BM2017.0099
|
| [227] |
Ye S, Shen F, Jiao L, et al. (2020) Biosynthesis of selenoproteins by Saccharomyces cerevisiae and characterization of its antioxidant activities. Int J Biol Macromol 164: 3438-3445. https://doi.org/10.1016/j.ijbiomac.2020.08.144
|
| [228] |
Babaei M, Borja Zamfir GM, Chen X, et al. (2020) Metabolic engineering of Saccharomyces cerevisiae for rosmarinic acid production. ACS Synth Biol 9: 1978-1988. https://doi.org/10.1021/acssynbio.0c00048
|
| [229] |
Callari R, Fischer D, Heider H, et al. (2018) Biosynthesis of angelyl-CoA in Saccharomyces cerevisiae. Microb Cell Factories 17: 72. https://doi.org/10.1186/s12934-018-0925-8
|
| [230] |
Sun S, Xu X, Liang L, et al. (2021) Lactic acid-producing probiotic Saccharomyces cerevisiae attenuates ulcerative colitis via suppressing macrophage pyroptosis and modulating gut microbiota. Front Immunol 12: 777665. https://doi.org/10.3389/fimmu.2021.777665
|
| [231] |
Hu Q, Yu L, Zhai Q, et al. (2023) Anti-inflammatory, barrier maintenance, and gut microbiome modulation effects of Saccharomyces cerevisiae QHNLD8L1 on dss-induced ulcerative colitis in mice. Int J Mol Sci 24: 6721. https://doi.org/10.3390/ijms24076721
|
| [232] | Fragopoulou E, Antonopoulou S, Tsoupras A, et al. Antiatherogenic properties of red/white wine, musts, grape-skins, and yeast, 25–29 (2004). Available from: https://scholar.google.com/scholar?hl=el&as_sdt=0%2C5&q=Fragopoulou%2C+E.%2C+Antonopoulou%2C+S.%2C+Tsoupras%2C+A.%2C+Tsantila%2C+N.%2C+Grypioti%2C+A.%2C+Gribilas%2C+G.%2C+Gritzapi%2C+H.%2C+Konsta%2C+E.%2C+Skandalou%2C+E.%2C+%26+Papadopoulou%2C+A.+%282004%29.+Antiatherogenic+properties+of+red%2Fwhite+wine%2C+musts%2C+grape-skins%2C+and+yeast.+25%E2%80%9329.&btnG= |
| [233] |
Csoma H, Acs-Szabo L, Papp LA, et al. (2023) Characterization of Zygosaccharomyces lentus yeast in hungarian botrytized wines. Microorganisms 11: 852. https://doi.org/10.3390/microorganisms11040852
|
| [234] |
Castro L, Gómez-Álvarez H, González F, et al. (2023) Biorecovery of rare earth elements from fluorescent lamp powder using the fungus Aspergillus niger in batch and semicontinuous systems. Miner Eng 201: 108215. https://doi.org/10.1016/j.mineng.2023.108215
|
| [235] |
Steels H, James SA, Roberts IN, et al. (1999) Zygosaccharomyces lentus: a significant new osmophilic, preservative-resistant spoilage yeast, capable of growth at low temperature: H. STEELS ET AL. J Appl Microbiol 87: 520-527. https://doi.org/10.1046/j.1365-2672.1999.00844.x
|
| [236] | Mcmeeking A, Dieckmann E, Cheeseman C (2024) Production methods for bacterial biomaterials: A review. Mater Today Sustain 25: 100623. https://doi.org/10.1016/j.mtsust.2023.100623 |
| [237] |
Stadie J, Gulitz A, Ehrmann MA, et al. (2013) Metabolic activity and symbiotic interactions of lactic acid bacteria and yeasts isolated from water kefir. Food Microbiol 35: 92-98. https://doi.org/10.1016/j.fm.2013.03.009
|
| [238] |
Gientka I, Kieliszek M, Jermacz K, et al. (2017) Identification and characterization of oleaginous yeast isolated from kefir and its ability to accumulate intracellular fats in deproteinated potato wastewater with different carbon sources. BioMed Res Int 2017: 1-19. https://doi.org/10.1155/2017/6061042
|
| [239] |
Lencioni L, Romani C, Gobbi M, et al. (2016) Controlled mixed fermentation at winery scale using Zygotorulaspora florentina and Saccharomyces cerevisiae. Int J Food Microbiol 234: 36-44. https://doi.org/10.1016/j.ijfoodmicro.2016.06.004
|
| [240] |
Hosseini M, Sharifan A (2021) Biological properties of yeast-based mannoprotein for prospective biomedical applications. Comb Chem High Throughput Screen 24: 831-840. https://doi.org/10.2174/1386207323999200818162030
|
| [241] | Alsayadi M, Al Jawfi Y, Belarbi M, et al. (2013) Antioxidant potency of water kefir. J Microbiol Biotechnol Food Sci 2: 2444-2447. Available from: https://office2.jmbfs.org/index.php/JMBFS/article/view/7101 |
| [242] | Constantin EA, Popa-Tudor I, Matei F, et al. Evaluation of polyphenol content and antioxidant activity of standard water kefir, NeXT-Chem 2023, MDPI, 7. (2023). https://doi.org/10.3390/chemproc2023013007 |
| [243] | Vamsnu E, Dangnon DB (2023) Characterizing water kefir beverages with antioxidant effects: preliminary analysis. Sci Bull Ser F Biotechnol 27. Available from: https://biotechnologyjournal.usamv.ro/pdf/2023/issue_2/Art12.pdf |
| [244] |
Güzel-Seydim ZB, Şatır G, Gökırmaklı Ç (2023) Use of mandarin and persimmon fruits in water kefir fermentation. Food Sci Nutr 11: 5890-5897. https://doi.org/10.1002/fsn3.3561
|
| [245] |
Falsoni RMP, Moraes FDSA, Rezende MSD, et al. (2022) Pretreatment with water kefir reduces the development of acidified ethanol-induced gastric ulcers. Braz J Pharm Sci 58: e191046. https://doi.org/10.1590/s2175-97902022e191046
|
| [246] |
Diniz RO, Perazzo FF, Carvalho JCT, et al. (2003) Atividade antiinflamatória de quefir, um probiótico da medicina popular. Rev Bras Farmacogn 13: 19-21. https://doi.org/10.1590/S0102-695X2003000300008
|
| [247] |
Rodrigues KL, Carvalho JCT, Schneedorf JM (2005) Anti-inflammatory properties of kefir and its polysaccharide extract. InflammoPharmacology 13: 485-492. https://doi.org/10.1163/156856005774649395
|
| [248] | Aligita W, Tarigan PN, Susilawati E (2020) Anti inflammatory and antioxidant activity of kefir water. Int J Biol Pharm Allied Sci 9. https://doi.org/10.31032/IJBPAS/2020/9.1.4904 |
| [249] | Aligita A, Si WM Anti inflammatory and antioxidant activity of kefir water (2022). Available from: https://scholar.google.com/scholar?hl=el&as_sdt=0%2C5&q=ALIGITA%2C+A.%2C+%26+Si%2C+W+M.+%282022%29.+Anti+inflammatory+and+antioxidant+activity+of+kefir+water.&btnG= |
| [250] |
Calatayud M, Börner RA, Ghyselinck J, et al. (2021) Water kefir and derived pasteurized beverages modulate gut microbiota, intestinal permeability and cytokine production In Vitro. Nutrients 13: 3897. https://doi.org/10.3390/nu13113897
|
| [251] |
Talib N, Mohamad NE, Yeap SK, et al. (2019) Isolation and characterization of Lactobacillus spp. from kefir samples in Malaysia. Molecules 24: 2606. https://doi.org/10.3390/molecules24142606
|
| [252] |
Gökırmaklı Ç, Erol Z, Gun I, et al. (2023) Prophylaxis effects of water kefir on post-infectious irritable bowel syndrome in rat model. Int J Food Sci Technol 58: 3371-3378. https://doi.org/10.1111/ijfs.16310
|
| [253] |
Guven M, Akman T, Yener AU, et al. (2015) The neuroprotective effect of kefir on spinal cord ischemia/reperfusion injury in rats. J Korean Neurosurg Soc 57: 335. https://doi.org/10.3340/jkns.2015.57.5.335
|
| [254] |
Yurtal Z, Kutlu T, Altuğ M, et al. (2022) Investigation of the neuroprotective effect of kefir in experimental spinal cord injury. Ank Üniversitesi Vet Fakültesi Derg 70: 9-19. https://doi.org/10.33988/auvfd.872947
|
| [255] |
Kumar M, Yeap S, Lee H, et al. (2021) Selected kefir water from Malaysia attenuates hydrogen peroxide-induced oxidative stress by upregulating endogenous antioxidant levels in SH-SY5Y neuroblastoma cells. Antioxidants 10: 940. https://doi.org/10.3390/antiox10060940
|
| [256] |
Alsayadi M, Jawfi YA, Belarbi M, et al. (2014) Evaluation of anti-hyperglycemic and anti-hyperlipidemic activities of water kefir as probiotic on streptozotocin-induced diabetic wistar rats. J Diabetes Mellit 04: 85-95. https://doi.org/10.4236/jdm.2014.42015
|
| [257] |
Rocha-Gomes A, Escobar A, Soares JS, et al. (2018) Chemical composition and hypocholesterolemic effect of milk kefir and water kefir in Wistar rats. Rev Nutr 31: 137-145. https://doi.org/10.1590/1678-98652018000200001
|
| [258] |
Aligita W, Singgih M, Sutrisno E, et al. (2023) Hepatoprotective study of Indonesian water kefir against CCl4-induced liver injury in rats. J Pharm Pharmacogn Res 11: 1002-1016. https://doi.org/10.56499/jppres23.1732_11.6.1002
|
| [259] |
Ye Z, Yang X, Deng B, et al. (2023) Prevention of DSS-induced colitis in mice with water kefir microbiota via anti-inflammatory and microbiota-balancing activity. Food Funct 14: 6813-6827. https://doi.org/10.1039/D3FO00354J
|
| [260] |
Moreira MEC, Santos MHD, Zolini GPP, et al. (2008) Anti-inflammatory and cicatrizing activities of a carbohydrate fraction isolated from sugary kefir. J Med Food 11: 356-361. https://doi.org/10.1089/jmf.2007.329
|
| [261] | Aligita W, Singgih M, Sutrisno E, et al. (2023) Protein-protein interaction analysis to identify nuclear factor-erythroid-2 factor 2 (nrf2) inhibition by extracellular enzymes from water kefir organisms. Int J Appl Pharm 109–112. https://doi.org/10.22159/ijap.2023.v15s2.20 |
| [262] |
Rodrigues KL, Caputo LRG, Carvalho JCT, et al. (2005) Antimicrobial and healing activity of kefir and kefiran extract. Int J Antimicrob Agents 25: 404-408. https://doi.org/10.1016/j.ijantimicag.2004.09.020
|
| [263] |
Brasil GA, Andrade Moraes FS, Prucoli Falsoni RM, et al. (2019) Pretreatment with water kefir promotes a decrease in ulcer development in an ethanol-acidified ulcer model. FASEB J 33: 760-762. https://doi.org/10.1096/fasebj.2019.33.1_supplement.760.2
|
| [264] |
Rodrigues KL, Araújo TH, Schneedorf JM, et al. (2016) A novel beer fermented by kefir enhances anti-inflammatory and anti-ulcerogenic activities found isolated in its constituents. J Funct Foods 21: 58-69. http://dx.doi.org/10.1016/j.jff.2015.11.035
|
| [265] |
Mechmeche M, Ksontini H, Hamdi M, et al. (2019) Production of bioactive peptides in tomato seed protein isolate fermented by water kefir culture: optimization of the fermentation conditions. Int J Pept Res Ther 25: 137-150. https://doi.org/10.1007/s10989-017-9655-8
|
| [266] |
Azi F, Tu C, Meng L, et al. (2021) Metabolite dynamics and phytochemistry of a soy whey-based beverage bio-transformed by water kefir consortium. Food Chem 342: 128225. https://doi.org/10.1016/j.foodchem.2020.128225
|
| [267] |
Ozcelik F, Akan E, Kinik O (2021) Use of Cornelian cherry, hawthorn, red plum, roseship and pomegranate juices in the production of water kefir beverages. Food Biosci 42: 101219. https://doi.org/10.1016/j.fbio.2021.101219
|
| [268] |
Bueno RS, Ressutte JB, Hata NNY, et al. (2021) Quality and shelf life assessment of a new beverage produced from water kefir grains and red pitaya. LWT 140: 110770. https://doi.org/10.1016/j.lwt.2020.110770
|
| [269] |
Alrosan M, Tan TC, Easa AM, et al. (2023) Evaluation of quality and protein structure of natural water kefir-fermented quinoa protein concentrates. Food Chem 404: 134614. https://doi.org/10.1016/j.foodchem.2022.134614
|
| [270] |
Şafak H, Gün İ, Tudor Kalit M, et al. (2023) Physico-chemical, microbiological and sensory properties of water kefir drinks produced from demineralized whey and dimrit and shiraz grape varieties. Foods 12: 1851. https://doi.org/10.3390/foods12091851
|
| [271] |
Wang X, Wang P (2023) Red beetroot juice fermented by water kefir grains: physicochemical, antioxidant profile and anticancer activity. Eur Food Res Technol 249: 939-950. https://doi.org/10.1007/s00217-022-04185-7
|
| [272] |
Islamiana D, Prabowo R, Pramaningtyas MD (2020) The effect of orange water kefir on malondialdehyde (MDA) level and superoxide dismutase (SOD) inhibition rate in kidney tissue of the hyperlipidemic rat (Rattus norvegicus). Atherosclerosis 315: e264. https://doi.org/10.1016/j.atherosclerosis.2020.10.833
|
| [273] | Aspiras BEE, Flores R, Pareja MC (2015) Hepatoprotective effect of Fermented Water Kefir on Sprague-Dawley rats (Rattus norvegicus) induced with sublethal dose of Acetaminophen. Int J Curr Sci 17: 18-28. Available from: https://d1wqtxts1xzle7.cloudfront.net/47094518/Bea_Eunice-libre.pdf?1467943173=&response-content-disposition=inline%3B+filename%3DHepatoprotective_effect_of_Fermented_Wat.pdf&Expires=1725024033&Signature=e4qEln3fmYio4UeuPM8Qpo4pUYFf7rD~NSZA7u2tTveBvZ6TFh1b-LDYrMQG2PI4mYiReptaTHXIcsHFYxRXQSH5HPsyP-ceiPiLdd2VI0RyYBbhjheFE-ZB6aqkyg7AbpVsvH-UFu6n-ith9vB5lh1SYPqPh88M8EshTAwzWZePMMrHnD4V057tNH2IeOYJalMhHT4PfhQhSsU4DRz6eZoxgTdS~KPYHtdtVu1ZUGmrTGMua90YB3uyOhLK4JjH3tZhTkEyVgGnyNnZYaaQrBEmx22BbgEpC6~xlp-OvKvHGu-MaIlbZ~Pnq03PxXxgUP3VTi8W8I65CGfVuRe-Mg__&Key-Pair-Id=APKAJLOHF5GGSLRBV4ZA |
| [274] | Tiss M, Souiy Z, Abdeljelil NB, et al. (2020) Fermented soy milk prepared using kefir grains prevents and ameliorates obesity, type 2 diabetes, hyperlipidemia and Liver-Kidney toxicities in HFFD-rats. J FunctFoods 67: 103869. https://doi.org/10.1016/j.jff.2020.103869 |
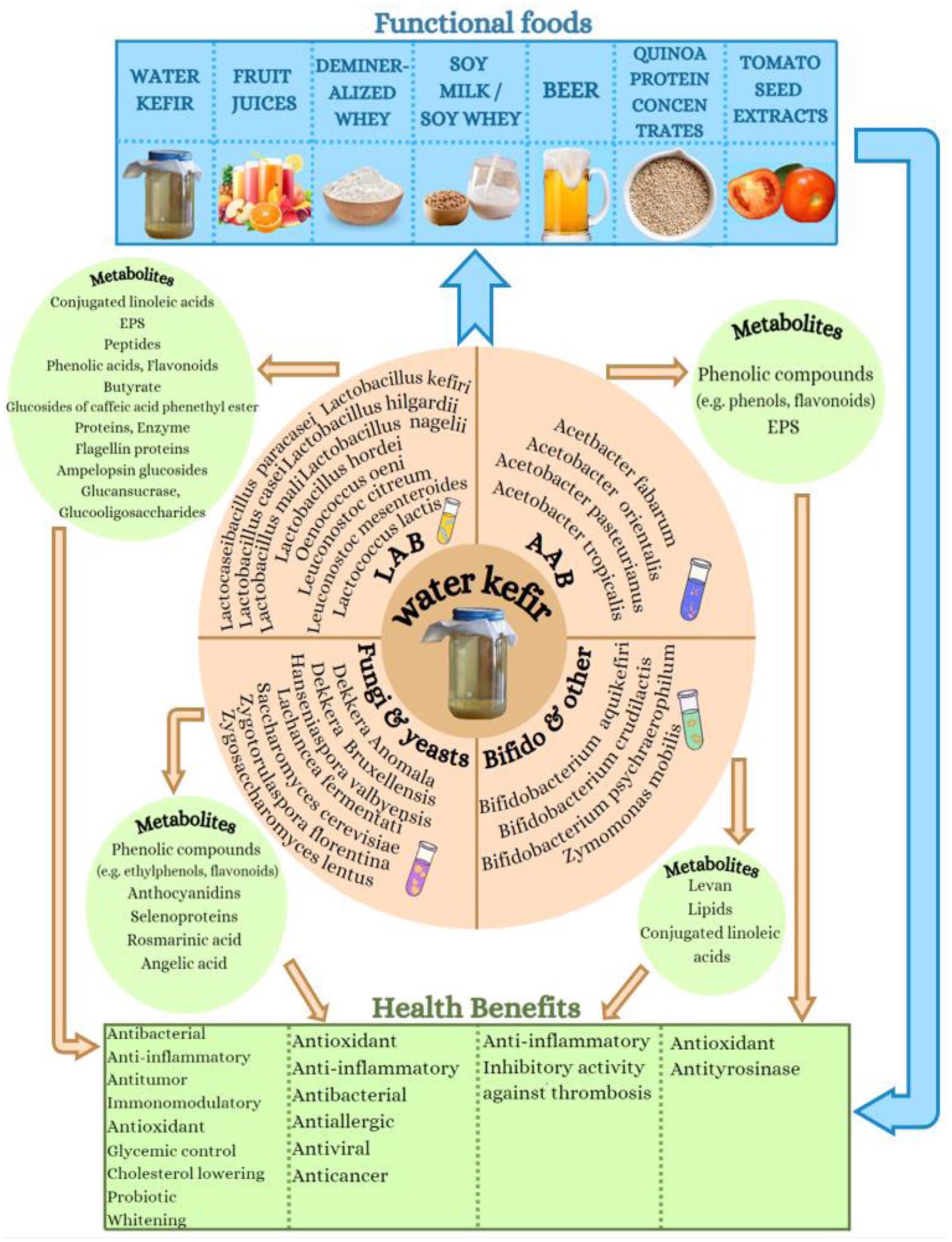
Figures(1) / Tables(6)

Dimitra Papadopoulou, Vasiliki Chrysikopoulou, Aikaterini Rampaouni, Alexandros Tsoupras. Antioxidant and anti-inflammatory properties of water kefir microbiota and its bioactive metabolites for health promoting bio-functional products and applications[J]. AIMS Microbiology, 2024, 10(4): 756-811. doi: 10.3934/microbiol.2024034







 DownLoad:
DownLoad: