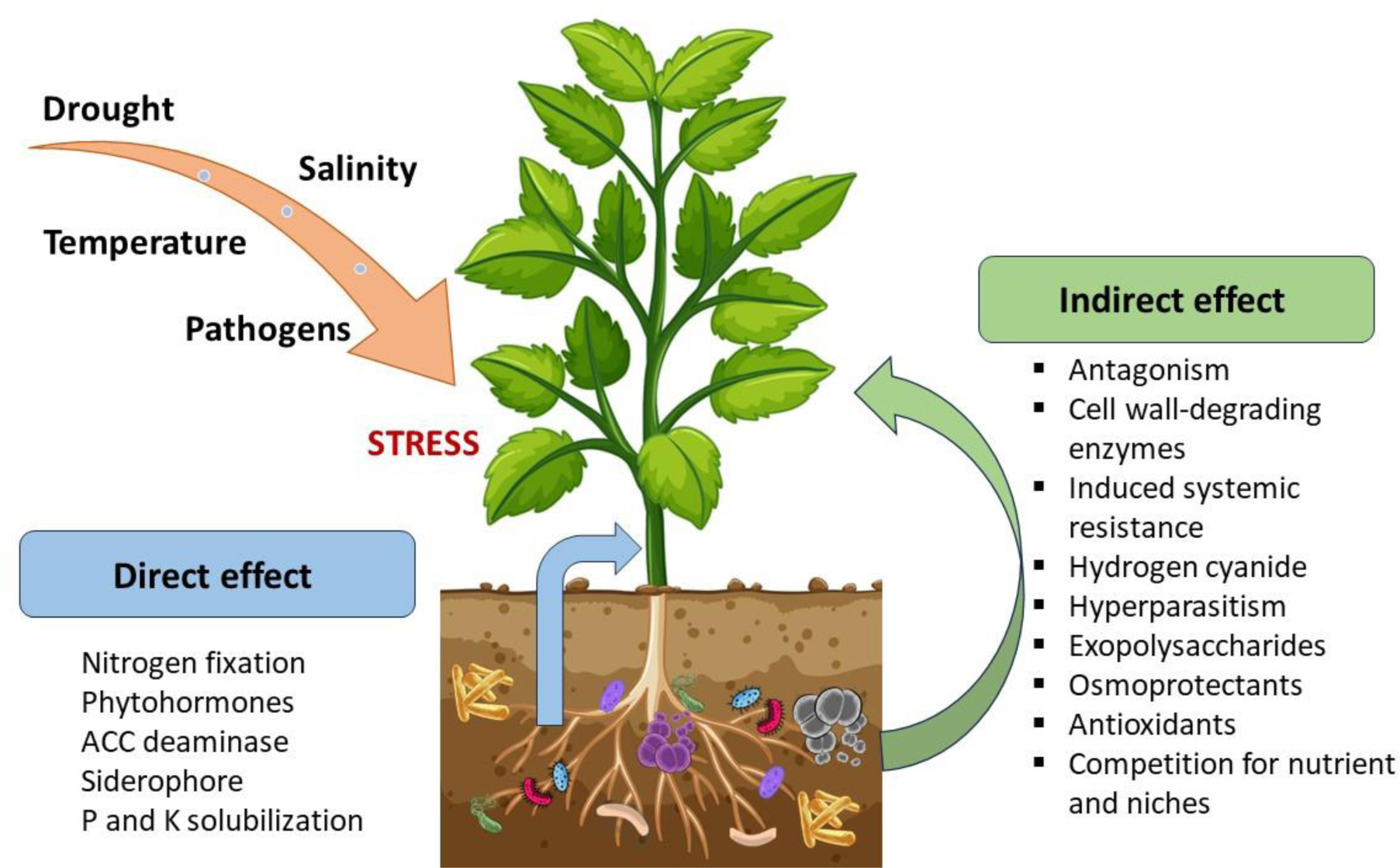
Endophytic bacteria live asymptomatically inside the tissues of host plants without inflicting any damage. Endophytes can confer several beneficial traits to plants, which can contribute to their growth, development, and overall health. They have been found to stimulate plant growth by enhancing nutrient uptake and availability. They can produce plant growth-promoting substances such as auxins, cytokinins, and gibberellins, which regulate various aspects of plant growth and development. Endophytes can also improve root system architecture, leading to increased nutrient and water absorption. Some endophytes possess the ability to solubilize nutrients, such as phosphorus and potassium, making them more available for plant uptake, and fixing atmospheric nitrogen. Chickpea (Cicer arietinum) is a major legume crop that has mutualistic interactions with endophytes. These endophytes can benefit the chickpea plant in various ways, including higher growth, improved nutrient uptake, increased tolerance to abiotic and biotic stressors, and disease suppression. They can produce enzymes and metabolites that scavenge harmful reactive oxygen species, thus reducing oxidative stress. Moreover, several studies reported that endophytes produce antimicrobial compounds, lytic enzymes, and volatile organic compounds that inhibit the growth of fungal pathogens and trigger systemic defense responses in plants, leading to increased resistance against a broad range of pathogens. They can activate plant defense pathways, including the production of defense-related enzymes, phytoalexins, and pathogenesis-related proteins, thereby providing long-lasting protection. It is important to note that the diversity and function of chickpea-associated endophytes can vary depending on factors such as variety, geographical location, and environmental conditions. The mechanisms behind the plant-beneficial interactions are still being intensively explored. In this review, new biotechnologies in agricultural production and ecosystem stability were presented. Thus, harnessing chickpea endophytes could be exploited in developing drought-resistant cultivars that can maintain productivity in arid and semi-arid environments, crucial for meeting the global demand for chickpeas.
Citation: Yulduzkhon Abdullaeva, Gulsanam Mardonova, Farkhod Eshboev, Massimiliano Cardinale, Dilfuza Egamberdieva. Harnessing chickpea bacterial endophytes for improved plant health and fitness[J]. AIMS Microbiology, 2024, 10(3): 489-506. doi: 10.3934/microbiol.2024024
Endophytic bacteria live asymptomatically inside the tissues of host plants without inflicting any damage. Endophytes can confer several beneficial traits to plants, which can contribute to their growth, development, and overall health. They have been found to stimulate plant growth by enhancing nutrient uptake and availability. They can produce plant growth-promoting substances such as auxins, cytokinins, and gibberellins, which regulate various aspects of plant growth and development. Endophytes can also improve root system architecture, leading to increased nutrient and water absorption. Some endophytes possess the ability to solubilize nutrients, such as phosphorus and potassium, making them more available for plant uptake, and fixing atmospheric nitrogen. Chickpea (Cicer arietinum) is a major legume crop that has mutualistic interactions with endophytes. These endophytes can benefit the chickpea plant in various ways, including higher growth, improved nutrient uptake, increased tolerance to abiotic and biotic stressors, and disease suppression. They can produce enzymes and metabolites that scavenge harmful reactive oxygen species, thus reducing oxidative stress. Moreover, several studies reported that endophytes produce antimicrobial compounds, lytic enzymes, and volatile organic compounds that inhibit the growth of fungal pathogens and trigger systemic defense responses in plants, leading to increased resistance against a broad range of pathogens. They can activate plant defense pathways, including the production of defense-related enzymes, phytoalexins, and pathogenesis-related proteins, thereby providing long-lasting protection. It is important to note that the diversity and function of chickpea-associated endophytes can vary depending on factors such as variety, geographical location, and environmental conditions. The mechanisms behind the plant-beneficial interactions are still being intensively explored. In this review, new biotechnologies in agricultural production and ecosystem stability were presented. Thus, harnessing chickpea endophytes could be exploited in developing drought-resistant cultivars that can maintain productivity in arid and semi-arid environments, crucial for meeting the global demand for chickpeas.

| [1] | Barrera VA, Martin ME, Aulicino M, et al. (2019) Carbon-substrate utilization profiles by Cladorrhinum (Ascomycota). Rev Argent Microbiol 51: 302-306. https://doi.org/10.1016/j.ram.2018.09.005 |
| [2] |
Bagy HMMK, Hassan EA, Nafady NA, et al. (2019) Efficacy of arbuscular mycorrhizal fungi and endophytic strain Epicoccum nigrum ASU11 as biocontrol agents against blackleg disease of potato caused by bacterial strain Pectobacterium carotovora subsp. atrosepticum PHY7. Biol Control 134: 103-113. https://doi.org/10.1016/j.biocontrol.2019.03.005 
|
| [3] |
Abaya A, Xue A, Hsiang T (2021) Selection and screening of fungal endophytes against wheat pathogens. Biol Control 154: 104511. https://doi.org/10.1016/j.biocontrol.2020.104511
|
| [4] |
Eshboev F, Egamberdieva D (2024) Medicinal plant-associated endophytic fungi: metabolites and bioactivity. Plant Endophytes and Secondary Metabolites . New York: Academic Press 95-104. https://doi.org/10.1016/B978-0-443-13365-7.00013-0
|
| [5] |
Rachwa-Rosiak D, Nebesny E, Budryn G (2015) Chickpeas—composition, nutritional value, health benefits, application to bread and snacks: A review. Crit Rev Food Sci Nutr 55: 1137-1145. https://doi.org/10.1080/10408398.2012.687418
|
| [6] |
Pushpavalli R, Berger JD, Turner NC, et al. (2020) Cross-tolerance for drought, heat and salinity stresses in chickpea (Cicer arietinum L.). J Agronomy Crop Science 206: 405-419. https://doi.org/10.1111/jac.12393
|
| [7] |
Romdhane SB, Trabelsi M, Aouani ME, et al. (2009) The diversity of rhizobia nodulating chickpea (Cicer arietinum) under water deficiency as a source of more efficient inoculants. Soil Biol Biochem 41: 2568-2572. https://doi.org/10.1016/j.soilbio.2009.09.020
|
| [8] |
Egamberdieva D, Li L, Ma H, et al. (2019) Soil amendment with different maize biochars improves chickpea growth under different moisture levels by improving symbiotic performance with Mesorhizobium ciceri and soil biochemical properties to varying degrees. Front Microbiol 10: 2423. https://doi.org/10.3389/fmicb.2019.02423
|
| [9] |
Qiu T, Peñuelas J, Chen Y, et al. (2024) Arbuscular mycorrhizal fungal interactions bridge the support of root-associated microbiota for slope multifunctionality in an erosion-prone ecosystem. iMeta 3: e187. https://doi.org/10.1002/imt2.187
|
| [10] |
Tedersoo L, Bahram M (2019) Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes. Biol Rev Camb Philos Soc 94: 1857-1880. https://doi.org/10.1111/brv.12538
|
| [11] | Ahmad P, Hashem A, Abd-Allah EF, et al. (2015) Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L) through antioxidative defense system. Front Plant Sci 6: 868. https://doi.org/10.3389/fpls.2015.00868 |
| [12] |
Ruiz-Lozano JM, Collados C, Barea JM, et al. (2001) Arbuscular mycorrhizal symbiosis can alleviate drought-induced nodule senescence in soybean plants. New Phytol 151: 493-502. https://doi.org/10.1046/j.0028-646x.2001.00196.x
|
| [13] | Egamberdieva D, Wirth S, Bellingrath-Kimura SD, et al. (2019) Salt-tolerant plant growth promoting rhizobacteria for enhancing crop productivity of saline soils. Front Microbiol 10: 469278. https://doi.org/10.3389/fmicb.2019.02791 |
| [14] |
Egamberdieva D, Shurigin V, Alaylar B, et al. (2020) The effect of biochars and endophytic bacteria on growth and root rot disease incidence of Fusarium infested narrow-leafed lupin (Lupinus angustifolius L). Microorganisms 8: 496. https://doi.org/10.3390/microorganisms8040496
|
| [15] |
Egamberdieva D, Wirth SJ, Alqarawi AA, et al. (2017) Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front Microbiol 8: 2104. https://doi.org/10.3389/fmicb.2017.02104
|
| [16] |
Cho ST, Chang HH, Egamberdieva D, et al. (2015) Genome analysis of Pseudomonas fluorescens PCL1751: A Rhizobacterium that controls root diseases and alleviates salt stress for its plant host. PLoS One 10: e0140231. https://doi.org/10.1371/journal.pone.0140231
|
| [17] |
Liu YH, Wei YY, Mohamad OAA, et al. (2019) Diversity, community distribution and growth promotion activities of endophytes associated with halophyte Lycium ruthenicum Murr. 3 Biotech 9: 144. https://doi.org/10.1007/s13205-019-1678-8
|
| [18] |
Mukherjee A, Singh BK, Verma JP (2020) Harnessing chickpea (Cicer arietinum L.) seed endophytes for enhancing plant growth attributes and bio-controlling against Fusarium sp. Microbiol Res 237: 126469. https://doi.org/10.1016/j.micres.2020.126469
|
| [19] |
Eshboev F, Karakozova M, Abdurakhmanov J, et al. (2023) Antimicrobial and cytotoxic activities of the secondary metabolites of endophytic fungi isolated from the medicinal plant Hyssopus officinalis. Antibiotics (Basel) 12: 1201. https://doi.org/10.3390/antibiotics12071201
|
| [20] |
Sessitsch A, Hardoim P, Döring J, et al. (2012) Functional characteristics of an endophyte community colonizing rice roots as revealed by metagenomic analysis. Mol Plant Microbe Interact 25: 28-36. https://doi.org/10.1094/MPMI-08-11-0204
|
| [21] |
Egamberdieva D, Kucharova Z (2009) Selection for root colonising bacteria stimulating wheat growth in saline soils. Biol Fertil Soils 45: 563-571. https://doi.org/10.1007/s00374-009-0366-y
|
| [22] |
Decunta FA, Pérez LI, Malinowski DP, et al. (2021) A systematic review on the effects of Epichloë fungal endophytes on drought tolerance in cool-season grasses. Front Plant Sci 12: 644731. https://doi.org/10.3389/fpls.2021.644731
|
| [23] |
Ben Gaied R, Sbissi I, Tarhouni M, et al. (2024) Bacterial endophytes from legumes native to arid environments are promising tools to improve Mesorhizobium-chickpea symbiosis under salinity. Biology 13: 96. https://doi.org/10.3390/biology13020096
|
| [24] | Rafique N, Khalil S, Cardinale M, et al. (2024) A comprehensive evaluation of the potential of plant growth-promoting rhizobacteria for applications in agriculture in stressed environments. Pedosphere . https://doi.org/10.1016/j.pedsph.2024.02.005 |
| [25] | Mukherjee A, Bhowmick S, Yadav S, et al. (2021) Re-vitalizing of endophytic microbes for soil health management and plant protection. 3 Biotech 11. https://doi.org/10.1007/s13205-021-02931-4 |
| [26] |
Brígido C, Singh S, Menéndez E, et al. (2019) Diversity and functionality of culturable endophytic bacterial communities in chickpea plants. Plants 8: 42. https://doi.org/10.3390/plants8020042
|
| [27] |
Glick BR (2014) Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol Res 169: 30-39. https://doi.org/10.1016/j.micres.2013.09.009
|
| [28] |
Berg G, Köberl M, Rybakova D, et al. (2017) Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol Ecol 93. https://doi.org/10.1093/femsec/fix050
|
| [29] |
Laranjeira SS, Alves IG, Marques G (2022) Chickpea (Cicer arietinum L.) seeds as a reservoir of endophytic plant growth-promoting bacteria. Curr Microbiol 79: 277. https://doi.org/10.1007/s00284-022-02942-1
|
| [30] |
Lalzar M, Zeevi A, Frenkel O, et al. (2022) Seed-derived microbial community of wild Cicer seedlings: Composition and augmentation to domesticated Cicer. Microbiol Spectr 10: e0278521. https://doi.org/10.1128/spectrum.02785-21
|
| [31] | Öğütçü H, Adiguzel A, Gulluce M, et al. (2009) Molecular characterization of Rhizobium strains isolated from wild chickpeas collected from high altitudes in Erzurum-Turkey. Rom Biotechnol Lett 14: 4294-4300. https://doi.org/10.1046/j.1365-2672.2002.01718.x |
| [32] |
Laranjo M, Alexandre A, Rivas R (2008) Chickpea rhizobia symbiosis genes are highly conserved across multiple Mesorhizobium species: Mesorhizobia symbiosis genes are conserved. FEMS Microbiol Ecol 66: 391-400. https://doi.org/10.1111/j.1574-6941.2008.00584.x
|
| [33] |
Alok D, Annapragada H, Singh S, et al. (2020) Symbiotic nitrogen fixation and endophytic bacterial community structure in Bt-transgenic chickpea (Cicer arietinum L). Sci Rep 10: 5453. https://doi.org/10.1038/s41598-020-62199-1
|
| [34] |
Ahmad M, Naseer I, Hussain A, et al. (2019) Appraising endophyte-plant symbiosis for improved growth, nodulation, nitrogen fixation and abiotic stress tolerance: An experimental investigation with chickpea (Cicer arietinum L.). Agronomy 9: 621. https://doi.org/10.3390/agronomy9100621
|
| [35] |
Misk A, Franco C (2011) Biocontrol of chickpea root rot using endophytic actinobacteria. Biocontrol (Dordrecht) 56: 811-822. https://doi.org/10.1007/s10526-011-9352-z
|
| [36] | Khanna A, Raj K, Kumar P, et al. (2022) Antagonistic and growth-promoting potential of multifarious bacterial endophytes against Fusarium wilt of chickpea. Egypt J Biol Pest Contr 32. https://doi.org/10.1186/s41938-022-00516-8 |
| [37] |
Akhtar MS, Siddiqui ZA (2009) Use of plant growth-promoting rhizobacteria for the biocontrol of root-rot disease complex of chickpea. Australasian Plant Pathol 38: 44-50. https://doi.org/10.1071/AP08075
|
| [38] |
Alexandre A, Brígido C, Laranjo M, et al. (2009) Survey of chickpea rhizobia diversity in Portugal reveals the predominance of species distinct from Mesorhizobium ciceri and Mesorhizobium mediterraneum. Microb Ecol 58: 930-941. https://doi.org/10.1007/s00248-009-9536-6
|
| [39] |
Brígido C, Glick BR, Oliveira S (2017) Survey of plant growth-promoting mechanisms in native Portuguese chickpea Mesorhizobium isolates. Microb Ecol 73: 900-915. https://doi.org/10.1007/s00248-016-0891-9
|
| [40] |
Egamberdieva D, Wirth SJ, Shurigin VV, et al. (2017) Endophytic bacteria improve plant growth, symbiotic performance of chickpea (Cicer arietinum L.) and induce suppression of root rot caused by fusarium solani under salt stress. Front Microbiol 8: 1887. https://doi.org/10.3389/fmicb.2017.01887
|
| [41] |
Vijayabharathi R, Gopalakrishnan S, Sathya A, et al. (2018) Deciphering the tri-dimensional effect of endophytic Streptomyces sp. on chickpea for plant growth promotion, helper effect with Mesorhizobium ciceri and host-plant resistance induction against Botrytis cinerea. Microb Pathog 122: 98-107. https://doi.org/10.1016/j.micpath.2018.06.019
|
| [42] |
Kushwaha P, Srivastava R, Pandiyan K, et al. (2021) Enhancement in plant growth and zinc biofortification of chickpea (Cicer arietinum L.) by Bacillus altitudinis. J Soil Sci Plant Nutr 21: 922-935. https://doi.org/10.1007/s42729-021-00411-5
|
| [43] |
Khan N, Bano A, Rahman MA, et al. (2019) Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci Rep 9: 2097. https://doi.org/10.1038/s41598-019-38702-8
|
| [44] |
Nautiyal CS (1997) Rhizosphere competence of Pseudomonas sp. NBRI9926 and Rhizobium sp. NBRI9513 involved in the suppression of chickpea (Cicer arietinum L.) pathogenic fungi. FEMS Microbiol Ecol 23: 145-158. https://doi.org/10.1111/j.1574-6941.1997.tb00398.x
|
| [45] |
Palmieri D, Vitullo D, De Curtis F, et al. (2017) A microbial consortium in the rhizosphere as a new biocontrol approach against fusarium decline of chickpea. Plant and soil 412: 425-439. https://doi.org/10.1007/s11104-016-3080-1
|
| [46] |
Maatallah J, Berraho EB, Munoz S, et al. (2002) Phenotypic and molecular characterization of chickpea rhizobia isolated from different areas of Morocco. J Appl Microbiol 93: 531-540. https://doi.org/10.1046/j.1365-2672.2002.01718.x
|
| [47] |
Landa BB, Hervás A, Bettiol W, et al. (1997) Antagonistic activity of bacteria from the chickpea rhizosphere against Fusarium Oxysporum f. sp. Ciceris. Phytoparasitica 25: 305-318. https://doi.org/10.1007/BF02981094
|
| [48] |
Truyens S, Weyens N, Cuypers A, et al. (2015) Bacterial seed endophytes: genera, vertical transmission and interaction with plants: Bacterial seed endophytes. Environ Microbiol Rep 7: 40-50. https://doi.org/10.1111/1758-2229.12181
|
| [49] |
Frank AC, Saldierna Guzmán JP, Shay JE (2017) Transmission of bacterial endophytes. Microorganisms 5: 70. https://doi.org/10.3390/microorganisms5040070
|
| [50] |
Johnson LJ, Johnson RD, Schardl CL, et al. (2003) Identification of differentially expressed genes in the mutualistic association of tall fescue with Neotyphodium coenophialum. Physiol Mol Plant Pathol 63: 305-317. https://doi.org/10.1016/j.pmpp.2004.04.001
|
| [51] |
Trivedi P, Leach JE, Tringe SG, et al. (2021) Author correction: Plant-microbiome interactions: from community assembly to plant health. Nat Rev Microbiol 19: 72. https://doi.org/10.1038/s41579-020-0412-1
|
| [52] |
Hardoim PR, van Overbeek LS, van Elsas JD (2008) Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol 16: 463-471. https://doi.org/10.1016/j.tim.2008.07.008
|
| [53] |
Vinagre F, Vargas C, Schwarcz K, et al. (2006) SHR5: a novel plant receptor kinase involved in plant–N2-fixing endophytic bacteria association. J Exp Bot 57: 559-569. https://doi.org/10.1093/jxb/erj041
|
| [54] |
Abdullaeva Y, Ratering S, Ambika Manirajan B, et al. (2022) Domestication impacts the wheat-associated microbiota and the rhizosphere colonization by seed-and soil-originated microbiomes, across different fields. Front Plant Sci 12. https://doi.org/10.3389/fpls.2021.806915
|
| [55] |
Gunnabo AH, van Heerwaarden J, Geurts R, et al. (2020) Symbiotic interactions between chickpea (Cicer arietinum L.) genotypes and Mesorhizobium strains. Symbiosis 82: 235-248. https://doi.org/10.1007/s13199-020-00724-6
|
| [56] |
Meyer D, Beuf D, Vekeman K (2015) A large diversity of non-rhizobial endophytes found in legume root nodules in Flanders (Belgium). Soil Biol Biochem 83: 1-11. https://doi.org/10.1016/j.soilbio.2015.01.002
|
| [57] |
Saini R, Adhikary A, Juneja S, et al. (2023) Drought priming triggers diverse metabolic adjustments and induces chilling tolerance in chickpea (Cicer arietinum L.). Plant Physiol Biochem 194: 418-439. https://doi.org/10.1016/j.plaphy.2022.11.034
|
| [58] | Mukherjee A, Gaurav A, Patel A, et al. (2021) Unlocking the potential plant growth-promoting properties of chickpea (Cicer arietinum L.) seed endophytes bio-inoculants for improving the soil health and crop production. Land Degrad Dev . https://doi.org/10.1002/ldr.4042 |
| [59] |
Bulgarelli D, Garrido-Oter R, Münch PC, et al. (2015) Structure and function of the bacterial root Microbiota in wild and domesticated barley. Cell Host Microbe 17: 392-403. https://doi.org/10.1016/j.chom.2015.01.011
|
| [60] |
Abdullaeva Y, Ambika Manirajan B, Honermeier B, et al. (2021) Domestication affects the composition, diversity, and co-occurrence of the cereal seed microbiota. J Adv Res 31: 75-86. https://doi.org/10.1016/j.jare.2020.12.008
|
| [61] |
Pérez-Jaramillo JE, Mendes R, Raaijmakers JM (2016) Impact of plant domestication on rhizosphere microbiome assembly and functions. Plant Mol Biol 90: 635-644. https://doi.org/10.1007/s11103-015-0337-7
|
| [62] |
Kim H, Lee KK, Jeon J, et al. (2020) Domestication of Oryza species eco-evolutionarily shapes bacterial and fungal communities in rice seed. Microbiome 8: 20. https://doi.org/10.1186/s40168-020-00805-0
|
| [63] |
Bazghaleh N, Hamel C, Gan Y, et al. (2018) Genotypic variation in the response of chickpea to arbuscular mycorrhizal fungi and non-mycorrhizal fungal endophytes. Canadian J Microbiol 64: 265-275. https://doi.org/10.1139/cjm-2017-0521
|
| [64] |
Khan N, Bano A, Zandi P (2018) Effects of exogenously applied plant growth regulators in combination with PGPR on the physiology and root growth of chickpea (Cicer arietinum) and their role in drought tolerance. J Plant Interac 13: 239-247. https://doi.org/10.1080/17429145.2018.1471527
|
| [65] |
Czarnes S, Hallett PD, Bengough AG, et al. (2000) Root-and microbial-derived mucilages affect soil structure and water transport. Eur J Soil Science 51: 435-443. https://doi.org/10.1046/j.1365-2389.2000.00327.x
|
| [66] |
Marasco R, Fusi M, Mosqueira M, et al. (2022) Rhizosheath–root system changes exopolysaccharide content but stabilizes bacterial community across contrasting seasons in a desert environment. Environ Microbiome 17. https://doi.org/10.1186/s40793-022-00407-3
|
| [67] |
Egamberdieva D, Wirth SJ, Shurigin VV, et al. (2017) Endophytic bacteria improve plant growth, symbiotic performance of chickpea (Cicer arietinum L.) and induce suppression of root rot caused by Fusarium solani under salt stress. Front Microbiol 8: 1887. https://doi.org/10.3389/fmicb.2017.01887
|
| [68] |
War AR, Paulraj MG, War MY (2011) Role of salicylic acid in induction of plant defense system in chickpea (Cicer arietinum L). Plant Signal Behav 6: 1787-1792. https://doi.org/10.4161/psb.6.11.17685
|
| [69] | Verma K, Saini R (2014) Recent advances in the regeneration and genetic transformation of soybean. J Innov Biol 1: 15-26. |
| [70] |
Chen J, Li Z, Xu D, et al. (2023) Patterns and drivers of microbiome in different rock surface soil under the volcanic extreme environment. iMeta 2: e122. https://doi.org/10.1002/imt2.122
|
| [71] | Caverzan A, Casassola A, Patussi Brammer S (2016) Reactive oxygen species and antioxidant enzymes involved in plant tolerance to stress. Abiotic and Biotic Stress in Plants-Recent Advances and Future Perspectives . London: IntechOpen. https://doi.org/10.5772/61368 |
| [72] | Noreen S, Ashraf M, Hussain M, et al. (2009) Exogenous application of salicylic acid enhances antioxidative capacity in salt stressed sunflower (Helianthus annuus L.) plants. Pakistan J Botany 41: 473-479. |
| [73] |
Bari R, Jones JDG (2009) Role of plant hormones in plant defence responses. Plant Mol Biol 69: 473-488. https://doi.org/10.1007/s11103-008-9435-0
|
| [74] |
Safdarian M, Askari H, Shariati JV, et al. (2019) Transcriptional responses of wheat roots inoculated with Arthrobacter nitroguajacolicus to salt stress. Sci Rep 9: 1792. https://doi.org/10.1038/s41598-018-38398-2
|
| [75] |
Suarez C, Cardinale M, Ratering S, et al. (2015) Plant growth-promoting effects of Hartmannibacter diazotrophicus on summer barley (Hordeum vulgare L.) under salt stress. Appl Soil Ecol 95: 23-30. https://doi.org/10.1016/j.apsoil.2015.04.017
|
| [76] |
Oliveira RS, Carvalho P, Marques G, et al. (2017) Increased protein content of chickpea (Cicer arietinum L.) inoculated with arbuscular mycorrhizal fungi and nitrogen-fixing bacteria under water deficit conditions. J Sci Food Agric 97: 4379-4385. https://doi.org/10.1002/jsfa.8201
|
| [77] |
Singh SP, Gaur R (2017) Endophytic Streptomyces spp. underscore induction of defense regulatory genes and confers resistance against Sclerotium rolfsii in chickpea. Biol Control 104: 44-56. https://doi.org/10.1016/j.biocontrol.2016.10.011
|
| [78] |
Salazar B, Ortiz A, Keswani C, et al. (2023) Bacillus spp. as bio-factories for antifungal secondary metabolites: Innovation beyond whole organism formulations. Microb Ecol 86: 1-24. https://doi.org/10.1007/s00248-022-02044-2
|
| [79] |
Šišić A, Baćanović-Šišić J, Al-Hatmi AMS, et al. (2018) The ‘forma specialis’ issue in Fusarium: A case study in Fusarium solani f. sp. pisi. Sci Rep 8. https://doi.org/10.1038/s41598-018-19779-z
|
| [80] | Fatima M, Ali SI, Thalho N, et al. (2021) Recent insight into production of cellulase by Fungi and its industrial applications. Ann Cell Biol 25: 1444-1472. Available from: http://annalsofrscb.ro/index.php/journal/article/view/10480 |
| [81] |
Mageshwaran V, Gupta R, Singh S, et al. (2022) Endophytic Bacillus subtilis antagonize soil-borne fungal pathogens and suppress wilt complex disease in chickpea plants (Cicer arietinum L.). Front Microbiol 13: 994847. https://doi.org/10.3389/fmicb.2022.994847
|
| [82] | Gorai PS, Ghosh R, Ghosh S, et al. (2023) Management of black root disease-causing fungus Fusarium solani CRP1 by endophytic Bacillus siamensis CNE6 through its metabolites and activation of plant defense genes. Microbiol Spectr e0308222. https://doi.org/10.1128/spectrum.03082-22 |
| [83] |
Sreevidya M, Gopalakrishnan S, Kudapa H, et al. (2016) Exploring plant growth-promotion actinomycetes from vermicompost and rhizosphere soil for yield enhancement in chickpea. Braz J Microbiol 47: 85-95. https://doi.org/10.1016/j.bjm.2015.11.030
|
| [84] |
AbdAllah E (2018) Endophytic bacterium Bacillus subtilis (BERA 71) improves salt tolerance in chickpea plants by regulating the plant defense mechanisms. J Plant Interact 13: 37-44. https://doi.org/10.1080/17429145.2017.1414321
|
| [85] | Zohary D, Hopf M (2000) Domestication of Plant in the Old World: The Origin and Spread of Cultivated Plants in West Asia, Europe and Nile Valley. Oxford: Oxford University Press. |
| [86] |
Paço A, Brígido C, Alexandre A, et al. (2016) The symbiotic performance of chickpea rhizobia can be improved by additional copies of the clpB chaperone gene. PLoS One 11: e0148221. https://doi.org/10.1371/journal.pone.0148221
|
| [87] |
Wanjofu EI, Venter SN, Beukes CW, et al. (2022) Nodulation and growth promotion of chickpea by Mesorhizobium isolates from diverse sources. Microorganisms 10: 2467. https://doi.org/10.3390/microorganisms10122467
|
| [88] |
Oke V, Long SR (1999) Bacteroid formation in the Rhizobium-legume symbiosis. Curr Opin Microbiol 2: 641-646. https://doi.org/10.1016/S1369-5274(99)00035-1
|
| [89] |
Moris M, Braeken K, Schoeters E, et al. (2005) Effective symbiosis between Rhizobium etli and Phaseolus vulgaris requires the alarmone ppGpp. J Bacteriol 187: 5460-5469. https://doi.org/10.1128/jb.187.15.5460-5469.2005
|
| [90] |
Calderón-Flores A, Pont D, Huerta-Saquero G (2005) The stringent response is required for amino acid and nitrate utilization, Nod factor regulation, nodulation, and nitrogen fixation in Rhizobium etli. J Bacteriol 187: 5075-5083. https://doi.org/10.1128/jb.187.15.5075-5083.2005
|
| [91] |
Wells DH, Long SR (2002) The Sinorhizobium meliloti stringent response affects multiple aspects of symbiosis: Stringent response in symbiosis. Mol Microbiol 43: 1115-1127. https://doi.org/10.1046/j.1365-2958.2002.02826.x
|
| [92] | Sepúlveda-Caamaño M, Gerding M, Vargas M, et al. (2018) Lentil (Lens culinaris L.) growth promoting rhizobacteria and their effect on nodulation in co inoculation with rhizobia. Arch Acker Pflanzenbau Bodenkd 64: 244-256. https://doi.org/10.1080/03650340.2017.1342034 |
Figures(1) / Tables(2)

Yulduzkhon Abdullaeva, Gulsanam Mardonova, Farkhod Eshboev, Massimiliano Cardinale, Dilfuza Egamberdieva. Harnessing chickpea bacterial endophytes for improved plant health and fitness[J]. AIMS Microbiology, 2024, 10(3): 489-506. doi: 10.3934/microbiol.2024024







 DownLoad:
DownLoad: