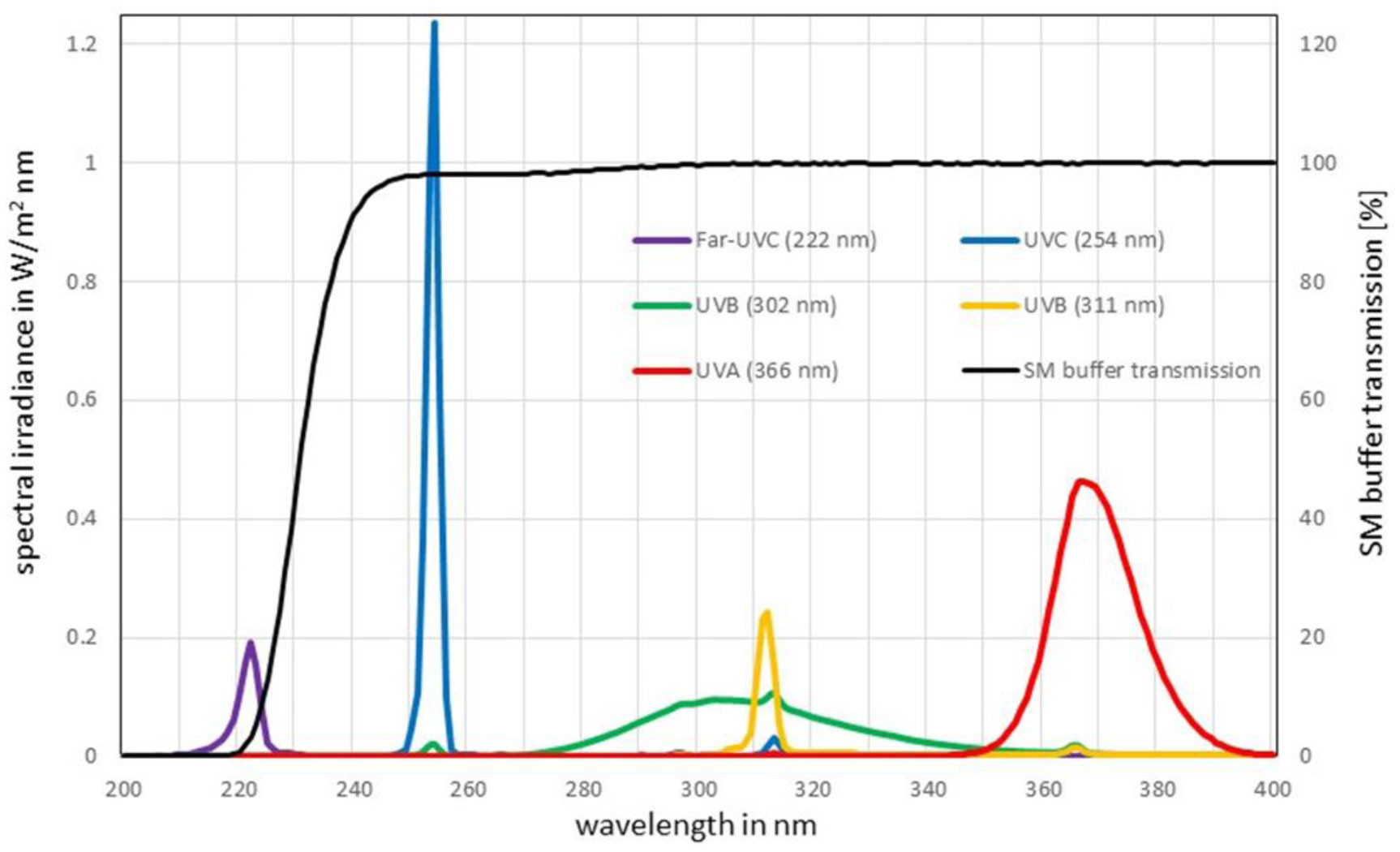
To minimize health risks, surrogates are often employed to reduce experiments with pathogenic microorganisms and the associated health risk. Due to structural similarities between the enveloped RNA-viruses SARS-CoV-2 and Phi6, the latter has been established as a nonpathogenic coronavirus surrogate for many applications. However, large discrepancies in the UV log-reduction doses between SARS-CoV-2 and Phi6 necessitate the search for a better surrogate for UV inactivation applications. A literature study provided the bacteriophage PhiX174 as a potentially more suitable nonpathogenic coronavirus surrogate candidate. In irradiation experiments, the sensitivity of PhiX174 was investigated upon exposure to UV radiation of wavelengths 222 nm (Far-UVC), 254 nm (UVC), 302 nm (broad-band UVB), 311 nm (narrow-band UVB) and 366 nm (UVA) using a plaque assay. The determined log-reduction doses for PhiX174 were 1.3 mJ/cm2 @ 222 nm, 5 mJ/cm2 @ 254 nm, 17.9 mJ/cm2 @ 302 nm, 625 mJ/cm2 @ 311 nm and 42.5 J/cm2 @ 366 nm. The comparison of these results with published log-reduction doses of SARS-CoV-2 in the same spectral region, led to the conclusion that the bacteriophage PhiX174 exhibits larger log-reduction doses than SARS-CoV-2, nevertheless, it is a better UV-surrogate at 222 nm (Far-UVC), 254 nm (UVC) and 302 nm (UVB) than the often applied Phi6.
Citation: Laura Weyersberg, Florian Sommerfeld, Petra Vatter, Martin Hessling. UV radiation sensitivity of bacteriophage PhiX174 - A potential surrogate for SARS-CoV-2 in terms of radiation inactivation[J]. AIMS Microbiology, 2023, 9(3): 431-443. doi: 10.3934/microbiol.2023023
To minimize health risks, surrogates are often employed to reduce experiments with pathogenic microorganisms and the associated health risk. Due to structural similarities between the enveloped RNA-viruses SARS-CoV-2 and Phi6, the latter has been established as a nonpathogenic coronavirus surrogate for many applications. However, large discrepancies in the UV log-reduction doses between SARS-CoV-2 and Phi6 necessitate the search for a better surrogate for UV inactivation applications. A literature study provided the bacteriophage PhiX174 as a potentially more suitable nonpathogenic coronavirus surrogate candidate. In irradiation experiments, the sensitivity of PhiX174 was investigated upon exposure to UV radiation of wavelengths 222 nm (Far-UVC), 254 nm (UVC), 302 nm (broad-band UVB), 311 nm (narrow-band UVB) and 366 nm (UVA) using a plaque assay. The determined log-reduction doses for PhiX174 were 1.3 mJ/cm2 @ 222 nm, 5 mJ/cm2 @ 254 nm, 17.9 mJ/cm2 @ 302 nm, 625 mJ/cm2 @ 311 nm and 42.5 J/cm2 @ 366 nm. The comparison of these results with published log-reduction doses of SARS-CoV-2 in the same spectral region, led to the conclusion that the bacteriophage PhiX174 exhibits larger log-reduction doses than SARS-CoV-2, nevertheless, it is a better UV-surrogate at 222 nm (Far-UVC), 254 nm (UVC) and 302 nm (UVB) than the often applied Phi6.

| [1] | Center for Systems Science and Engineering at Johns Hopkins University (Johns Hopkins University & Medicine Coronavirus Resource Center), COVID-19 Dashboard (2023). Available from: https://coronavirus.jhu.edu/map.html. |
| [2] | Gerhardt J (2022) Infektionsschutzgesetz (IfSG): Kommentar. München: C.H. Beck. |
| [3] | Robert Koch Institut, Zoonotischen Influenza bei Menschen (2023). Available from: https://www.rki.de/SharedDocs/FAQ/Gefluegelpest/Gefluegelpest.html. |
| [4] | Robert Koch Institut, Internationaler Mpox-Ausbruch: Einschätzung der Situation in Deutschland (2023). Available from: https://www.rki.de/DE/Content/InfAZ/A/Affenpocken/Ausbruch-2022-Situation-Deutschland.html?nn=2444038. |
| [5] |
Singh D, Soorneedi AR, Vaze N, et al. (2023) Assessment of SARS-CoV-2 surrogate inactivation on surfaces and in air using UV and blue light-based intervention technologies. J Air Waste Manag Assoc 73: 200-211. https://doi.org/10.1080/10962247.2022.2157907 
|
| [6] | Heßling M, Hönes K, Vatter P, et al. (2020) Ultraviolet irradiation doses for coronavirus inactivation - review and analysis of coronavirus photoinactivation studies. GMS Hyg Infect Control 15. https://doi.org/10.3205/dgkh000343 |
| [7] |
Seyer A, Sanlidag T (2020) Solar ultraviolet radiation sensitivity of SARS-CoV-2. Lancet Microbe 1: E8-E9. https://doi.org/10.1016/S2666-5247(20)30013-6
|
| [8] |
Kowalski W (2009) Ultraviolet germicidal irradiation handbook: UVGI for air and surface disinfection. Berlin: Springer.
|
| [9] |
Buonanno M, Welch D, Shuryak I (2020) Far-UVC light (222 nm) efficiently and safely inactivates airborne human coronaviruses. Sci Rep 10. https://doi.org/10.1038/s41598-020-67211-2
|
| [10] |
Zwicker P, Schleusener J, Lohan SB, et al. (2022) Application of 233 nm far-UVC LEDs for eradication of MRSA and MSSA and risk assessment on skin models. Sci Rep 12: 2587. https://doi.org/10.1038/s41598-022-06397-z
|
| [11] |
Buonanno M, Welch D, Brenner DJ (2021) Exposure of human skin models to KrCl excimer lamps: the impact of optical filtering. Photochem Photobiol 97: 517-523. https://doi.org/10.1111/php.13383
|
| [12] |
Welch D, Aquino de Muro M, Buonanno M, et al. (2022) Wavelength-dependent DNA photodamage in a 3-D human skin model over the far-UVC and germicidal UVC wavelength ranges from 215 to 255 nm. Photochem Photobiol 98: 1167-1171. https://doi.org/10.1111/PHP.13602
|
| [13] |
Welch D, Kleiman NJ, Arden PC, et al. (2023) No evidence of induced skin cancer or other skin abnormalities after long-term (66 week) chronic exposure to 222-nm far-UVC radiation. Photochem Photobiol 99: 168-175. https://doi.org/10.1111/php.13656
|
| [14] |
Eadie E, Barnard IMR, Ibbotson SH, et al. (2021) Extreme exposure to filtered far-UVC: a case study. Photochem Photobiol 97: 527-531. https://doi.org/10.1111/php.13385
|
| [15] |
Hickerson RP, Conneely MJ, Hirata Tsutsumi SK, et al. (2021) Minimal, superficial DNA damage in human skin from filtered far-ultraviolet C. Br J Dermatol 184: 1197-1199. https://doi.org/10.1111/bjd.19816
|
| [16] |
Boegel SJ, Gabriel M, Sasges M, et al. (2021) Robust evaluation of ultraviolet-C sensitivity for SARS-CoV-2 and surrogate coronaviruses. Microbiol Spectr 9: e0053721. https://doi.org/10.1128/Spectrum.00537-21
|
| [17] |
Normile D (2004) Mounting lab accidents raise SARS fears. Science 304: 659-661. https://doi.org/10.1126/science.304.5671.659
|
| [18] |
Della-Porta T (2008) Laboratory accidents and breaches in biosafety – they do occur!. Microbiol Aust 29: 62. https://doi.org/10.1071/MA08062
|
| [19] |
Cadnum JL, Li DF, Jones LD, et al. (2020) Evaluation of ultraviolet-C light for rapid decontamination of airport security bins in the era of SARS-CoV-2. Pathog Immun 5: 133-142. https://doi.org/10.20411/pai.v5i1.373
|
| [20] |
Prussin AJ, Schwake DO, Lin K, et al. (2018) Survival of the enveloped virus Phi6 in droplets as a function of relative humidity, absolute humidity, and temperature. Appl Environ Microbiol 84. https://doi.org/10.1128/aem.00551-18
|
| [21] |
Casanova LM, Weaver SR (2015) Evaluation of eluents for the recovery of an enveloped virus from hands by whole-hand sampling. J Appl Microbiol 118: 1210-1216. https://doi.org/10.1111/jam.12777
|
| [22] |
Ye Y, Ellenberg RM, Graham KE, et al. (2016) Survivability, partitioning, and recovery of enveloped viruses in untreated municipal wastewater. Environ Sci Technol 50: 5077-5085. https://doi.org/10.1021/acs.est.6b00876
|
| [23] |
Ma B, Gundy PM, Gerba CP, et al. (2021) UV inactivation of SARS-CoV-2 across the UVC spectrum: KrCl* excimer, mercury-vapor, and light-emitting-diode (LED) sources. Appl Environ Microbiol 87: e0153221. https://doi.org/10.1128/AEM.01532-21
|
| [24] |
Silverman AI, Boehm AB (2020) Systematic review and meta-analysis of the persistence and disinfection of human coronaviruses and their viral surrogates in water and wastewater. Environ Sci Technol Lett 7: 544-553. https://doi.org/10.1021/acs.estlett.0c00313
|
| [25] |
Aquino de Carvalho N, Stachler EN, Cimabue N, et al. (2017) Evaluation of Phi6 persistence and suitability as an enveloped virus surrogate. Environ Sci Technol 51: 8692-8700. https://doi.org/10.1021/acs.est.7b01296
|
| [26] |
Whitworth C, Mu Y, Houston H, et al. (2020) Persistence of bacteriophage phi 6 on porous and nonporous surfaces and the potential for its use as an Ebola virus or coronavirus surrogate. Appl Environ Microbiol 86. https://doi.org/10.1128/AEM.01482-20
|
| [27] |
Lytle CD, Budacz AP, Keville E, et al. (1991) Differential inactivation of surrogate viruses with merocyanine 540. Photochem Photobiol 54: 489-493. https://doi.org/10.1111/j.1751-1097.1991.tb02047.x
|
| [28] |
Costa L, Faustino MAF, Neves MGPMS, et al. (2012) Photodynamic inactivation of mammalian viruses and bacteriophages. Viruses 4: 1034-1074. https://doi.org/10.3390/v4071034
|
| [29] |
Weyersberg L, Klemens E, Buehler J, et al. (2022) UVC, UVB and UVA susceptibility of Phi6 and its suitability as a SARS-CoV-2 surrogate. AIMS Microbiol 8: 278-291. https://doi.org/10.3934/microbiol.2022020
|
| [30] | Sambrook J, Russell DW (2001) Molecular Cloning: A Laboratory Manual. Q Rev Biol 76: 348-349. https://doi.org/10.1086/394015 |
| [31] |
Cormier J, Janes M (2014) A double layer plaque assay using spread plate technique for enumeration of bacteriophage MS2. J Virol Methods 196: 86-92. https://doi.org/10.1016/j.jviromet.2013.10.034
|
| [32] |
Vatter P, Hoenes K, Hessling M (2021) Blue light inactivation of the enveloped RNA virus Phi6. BMC Res Notes 14: 187. https://doi.org/10.1186/s13104-021-05602-y
|
| [33] |
Setlow R, Boyce R (1960) The ultraviolet light inactivation of ΦX174 bacteriophage at different wave lengths and pH's. Biophys J 1: 29-41. https://doi.org/10.1016/S0006-3495(60)86873-7
|
| [34] | David CN (1964) UV inactivation and thymine dimerization in bacteriophage phi x. Z Vererbungsl 95: 318-325. https://doi.org/10.1007/BF01268664 |
| [35] |
Giese N, Darby J (2000) Sensitivity of microorganisms to different wavelengths of UV light: implications on modeling of medium pressure UV systems. Water Res 34: 4007-4013. https://doi.org/10.1016/S0043-1354(00)00172-X
|
| [36] |
Yarus M, Sinsheimer RL (1964) The UV-resistance of double-stranded PhiX174 DNA. J Mol Biol 8: 614-615. https://doi.org/10.1016/s0022-2836(64)80018-8
|
| [37] |
Rodriguez RA, Bounty S, Beck S, et al. (2014) Photoreactivation of bacteriophages after UV disinfection: role of genome structure and impacts of UV source. Water Res 55: 143-149. https://doi.org/10.1016/j.watres.2014.01.065
|
| [38] |
Battigelli DA, Sobsey MD, Lobe DC (1993) The inactivation of hepatitis a virus and other model viruses by UV irradiation. Water Sci Technol 27: 339-342. https://doi.org/10.2166/wst.1993.0371
|
| [39] |
Sommer R, Haider T, Cabaj A, et al. (1998) Time dose reciprocity in UV disinfection of water. Water Sci Technol 38: 145-150. https://doi.org/10.1016/S0273-1223(98)00816-6
|
| [40] |
Zuo X, Chu X, Hu J (2015) Effects of water matrix on virus inactivation using common virucidal techniques for condensate urine disinfection. Chemosphere 136: 118-124. https://doi.org/10.1016/j.chemosphere.2015.04.083
|
| [41] |
Sommer R (2001) Inactivation of bacteriophages in water by means of non-ionizing (uv-253.7nm) and ionizing (gamma) radiation: a comparative approach. Water Res 35: 3109-3116. https://doi.org/10.1016/S0043-1354(01)00030-6
|
| [42] |
Ho J, Seidel M, Niessner R, et al. (2016) Long amplicon (LA)-qPCR for the discrimination of infectious and noninfectious phix174 bacteriophages after UV inactivation. Water Res 103: 141-148. https://doi.org/10.1016/j.watres.2016.07.032
|
| [43] |
Proctor WR, Cook JS, Tennant RW (1972) Ultraviolet photobiology of Kilham rat virus and the absolute ultraviolet photosensitivities of other animal viruses: influence of DNA strandedness, molecular weight, and host-cell repair. Virology 49: 368-378. https://doi.org/10.1016/0042-6822(72)90489-8
|
| [44] |
Lee HS, Sobsey MD (2011) Survival of prototype strains of somatic coliphage families in environmental waters and when exposed to UV low-pressure monochromatic radiation or heat. Water Res 45: 3723-3734. https://doi.org/10.1016/j.watres.2011.04.024
|
| [45] |
Nuanualsuwan S, Mariam T, Himathongkham S, et al. (2002) Ultraviolet inactivation of feline calicivirus, human enteric viruses and coliphages. Photochem Photobiol 76: 406-410. https://doi.org/10.1562/0031-8655(2002)0760406UIOFCH2.0.CO2
|






Figures(7) / Tables(1)

Laura Weyersberg, Florian Sommerfeld, Petra Vatter, Martin Hessling. UV radiation sensitivity of bacteriophage PhiX174 - A potential surrogate for SARS-CoV-2 in terms of radiation inactivation[J]. AIMS Microbiology, 2023, 9(3): 431-443. doi: 10.3934/microbiol.2023023







 DownLoad:
DownLoad: