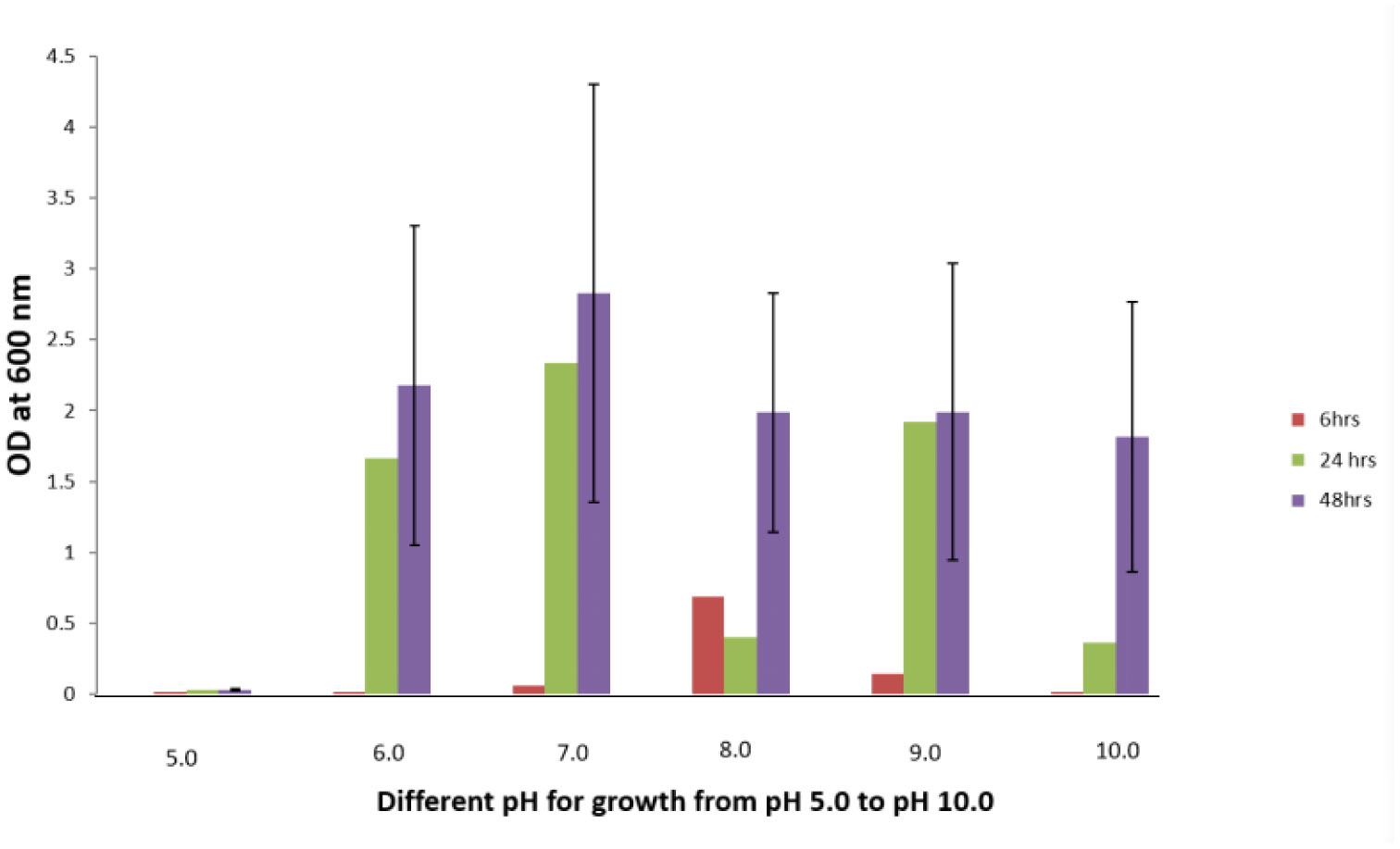
A Gram positive rod shaped bacterium designated as isolate H1 with Fluoride resistance up to 4 g/L sodium fluoride (NaF) in LB (Luria-Bertani) agar was isolated from a ground water sample of Narketpally area, Nalgonda district, Telangana, India. The colonies of isolate H1 were off white in color. Growth patterns of isolate H1 were observed at two different concentrations, 100 and 250 ppm, of NaF and also without NaF in the medium. In cases where NaF was present in the media, the lag phases of the growth curves were extended when compared to the absence of NaF. Optimum pH required for the organism's growth was 8. Isolate H1 required a temperature of 37 °C with 150 rpm and 2% NaCl for its optimal growth in the medium without NaF. Meanwhile, isolate H1 could thrive in a diverse pH range, i.e., pH 5–10, and at an NaCl concentration of up to 11% in the medium with NaF. Based on morphological, biochemical and molecular characterization, isolate H1 was identified as belonging to the genus Bacillus. It showed 98.47% 16S rDNA gene sequence similarity with Bacillus australimaris NH71_1T. Isolate H1 showed high fluoride removals of 22.5% and 38.2% with 100 and 250 mg/L of NaF in the LB broth when incubated at pH 8 and a temperature of 37 °C with 150 rpm for 3 day. Hence, this organism could be a promising isolate to apply for defluoridation of ground water in fluoride contaminthe ated areas.
Citation: M Thirumala, E Sai Krishna, P Sindhu Priya, S Vishnuvardhan Reddy. Characterization of a novel Fluoride resistant bacterial isolate and its capability of Fluoride bioremediation[J]. AIMS Microbiology, 2022, 8(4): 470-483. doi: 10.3934/microbiol.2022031
A Gram positive rod shaped bacterium designated as isolate H1 with Fluoride resistance up to 4 g/L sodium fluoride (NaF) in LB (Luria-Bertani) agar was isolated from a ground water sample of Narketpally area, Nalgonda district, Telangana, India. The colonies of isolate H1 were off white in color. Growth patterns of isolate H1 were observed at two different concentrations, 100 and 250 ppm, of NaF and also without NaF in the medium. In cases where NaF was present in the media, the lag phases of the growth curves were extended when compared to the absence of NaF. Optimum pH required for the organism's growth was 8. Isolate H1 required a temperature of 37 °C with 150 rpm and 2% NaCl for its optimal growth in the medium without NaF. Meanwhile, isolate H1 could thrive in a diverse pH range, i.e., pH 5–10, and at an NaCl concentration of up to 11% in the medium with NaF. Based on morphological, biochemical and molecular characterization, isolate H1 was identified as belonging to the genus Bacillus. It showed 98.47% 16S rDNA gene sequence similarity with Bacillus australimaris NH71_1T. Isolate H1 showed high fluoride removals of 22.5% and 38.2% with 100 and 250 mg/L of NaF in the LB broth when incubated at pH 8 and a temperature of 37 °C with 150 rpm for 3 day. Hence, this organism could be a promising isolate to apply for defluoridation of ground water in fluoride contaminthe ated areas.

| [1] |
Ghosh A, Mukherjee K, Ghosh SK, et al. (2013) Sources and toxicity of fluoride in the environment. Res Chem Intermed 39: 2881-2915. https://doi.org/10.1007/s11164-012-0841-1.2 
|
| [2] | (2006) WHOGuidelines for drinking-water quality. Recommendations incorporating the first and second addenda . Geneva: WHO 375-377. Available from: https://www.who.int/publications/i/item/97892415476113. |
| [3] |
Ying L, Jingmei Y, Bernd WB, et al. (2018) Genetic loci associated with fluoride resistance in Streptococcus mutans. Front Microbiol 9: 3093. https://doi.org/10.3389/fmicb.2018.03093
|
| [4] |
Maltz M, Emilson CG (1982) Susceptibility of oral bacteria to various fluoride salts. J Dent Res 61: 786-790. https://doi.org/10.1177/00220345820610062701
|
| [5] |
Juwarkar AA, Yadav SK (2010) Bioaccumulation and biotransformation of heavy metals. Bioremediation Technology . Amsterdam: Springer 266-284. https://link.springer.com/chapter/10.1007/978-90-481-3678-0_9
|
| [6] |
Curran TM, Buckley DH, Marquis RE (1994) Quasi-irreversible inhibition of enolase of Streptococcus mutans by fluoride. FEMS Microbiol Lett 119: 283-288. https://doi.org/10.1111/j.1574-6968.1994.tb06902.x
|
| [7] |
Guha CN, Clark AG, Sissons CH (1997) Inhibition of purified enolases from oral bacteria by fluoride. Oral Microbiol Immunol 12: 91-97. https://onlinelibrary.wiley.com/doi/10.1111/j.1399-302X.1997.tb00623.x
|
| [8] |
Burne RA, Marquis RE (2000) Alkali production by oral bacteria and protection against dental caries. FEMS Microbiol Lett 193: 1-6. https://doi.org/10.1111/j.1574-6968.2000.tb09393.x
|
| [9] |
van Loveren C, Hoogenkamp MA, Deng DM, et al. (2008) Effects of different kinds of fluorides on enolase and ATPase activity of a fluoride-sensitive and fluoride-resistant Streptococcus mutans strain. Caries Res 42: 429-434. https://doi.org/10.1159/000159606
|
| [10] |
Pandit S, Kim HJ, Song KY, et al. (2013) Relationship between fluoride concentration and activity against virulence factors and viability of a cariogenic biofilm: in vitro study. Caries Res 47: 539-547. https://doi.org/10.1159/000348519
|
| [11] | Sree KK, Raja CE, Ramesh U (2018) Isolation and characterization of fluoride resistant bacteria from groundwaters in Dindigul, Tamilnadu, India. Environ Res Technol 1: 69-74. https://dergipark.org.tr/en/download/article-file/460255 |
| [12] |
Shalaby MG, Al-Hossainy AF, Abo-Zeid AM, et al. (2022) Geotrichum candidum mediated [Cu8O7+P2O5] nanocomposite biofabrication, characterization, physicochemical properties and its in-vitro biocompatibility evaluation. J Inorg Organomet Polym 32: 2398-2415. https://doi.org/10.1007/s10904-022-02252-w
|
| [13] |
Hou HY, Li SY, Meng ZC, et al. (2021) Removal of Cu ions in wastewater through a combined foam separation cell adsorption approach. Chem Eng Tech 45: 585-594. https://doi.org/10.1002/ceat.202100482
|
| [14] | Darwesha OM, Shalaby MG, Abo-Zeid AM, et al. (2021) Nano-bioremediation of municipal wastewater using myco-synthesized iron nanoparticles. Egypt J Chem 64: 2499-2507. |
| [15] | Darwesh OM, Abd El-Latief AH, Abuarab ME, et al. (2021) Enhancing the efficiency of some agricultural wastes as low-cost absorbents to remove textile dyes from their contaminated solutions. Biomass Conv Bioref . https://doi.org/10.1007/s13399-020-01142-w |
| [16] |
Chouhan S, Tuteja U, Flora SJS (2012) Isolation, identification and characterization of fluoride resistant bacteria: possible role in bioremediation. Appl Biochem Microbiol 48: 43-50. https://doi.org/10.1134/S0003683812010036
|
| [17] |
Mukherjee S, Yadav V, Mondal M, et al. (2017) Characterization of a fluoride-resistant bacterium Acinetobacter sp. RH5 towards assessment of its water defluoridation capability. Appl Water Sci 7: 1923-1930. https://doi.org/10.1007/s13201-015-0370-3
|
| [18] |
Dutta M, Pan B, Ghosh K, et al. (2020) Isolation of Fluoride tolerant Bacillus spp (KT201599, KT201600) from the midgut of Drosophila melanogaster: Their probable role in fluoride removal. Proc Zool Soc 73: 175-183. https://doi.org/10.1007/s12595-018-0282-y
|
| [19] | Goutam B, Archya S, Tathagato R, et al. (2016) Isolation and characterization of fluoride resistant bacterial strains from fluoride endemic areas of west bengal, India: Assessment of their fluoride absorption efficiency. Res Report Fluoride 49: 429-440. |
| [20] |
Pal KC, Mondal NK, Chatterjee S, et al. (2014) Characterization of fluoride-tolerant halophilic Bacillus flexus NM25 (HQ875778) isolated from fluoride-affected soil in Birbhum district, West Bengal, India. Environ Monit Assess 186: 699-709. https://doi.org/10.1007/s10661-013-3408-8
|
| [21] |
Shiva SA, Dasaiah S, Pavan KP (2020) A study on bioremediation of fluoride-contaminated water via a novel bacterium Acinetobacter sp. (GU566361) isolated from potable water. Results Chem 2: 100070. https://doi.org/10.1016/j.rechem.2020.100070
|
| [22] |
Ying L, Bernd WB, Jiyao Li, et al. (2017) Fluoride resistance in Streptococcus mutans: a mini review. J Oral Microbiol 9: 1344509. https://doi.org/10.1080/20002297.2017.1344509
|
| [23] |
Brussock SM, Kral TA (1987) Effects of pH on expression of sodium fluoride resistance in Streptococcus mutans. J Dent Res 66: 1594-1596. https://doi.org/10.1177/00220345870660101701
|
| [24] |
Liao Y, Chen J, Brandt BW, et al. (2015) Identification and functional analysis of genome mutations in a fluoride-resistant Streptococcus mutans strain. Plos One 10: e0122630. https://doi.org/10.1371/journal.pone.0122630
|
| [25] |
Mitsuhata C, Puteri MM, Ohara Y, et al. (2014) Possible involvement of enolase in fluoride resistance in Streptococcus mutans. Pediatr Dental J 24: 12-16. https://doi.org/10.1016/j.pdj.2013.10.002
|
| [26] |
Zhang C, Zhang W, Xu J (2013) Isolation and Identification of methanethiol-utilizing bacterium CZo5 and its application in biotrickling filter of biogas. Bioresour Technol 150: 338-343. https://doi.org/10.1016/j.biortech.2013.10.024
|
| [27] | MacFaddin JF (1980) Biochemical tests for identification of medical bacteria. Philadelphia: Lippincott Williams & Wilkins. |
| [28] |
Evans JB, Kloos EW (1972) Use of shake cultures in a semisolid thioglycolate medium for differentiating staphylococci from micrococci. Appl Microbiol 23: 326-331. https://doi.org/10.1128/am.23.2.326-331.1972
|
| [29] |
Johnson JS, Spakowicz DJ, Hong B (2019) Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat Commun 10: 5029. https://doi.org/10.1038/s41467-019-13036-1
|
| [30] |
Peker N, Garcia-Croes S, Dijkhuizen B, et al. (2019) A comparison of three different bioinformatics analyses of the 16S–23S rRNA encoding region for bacterial identification. Front Microbiol 10: 620. https://doi.org/10.3389/fmicb.2019.00620
|
| [31] |
Chen YL, Lee CC, Lin YL, et al. (2015) Obtaining long 16S rDNA sequences using multiple primers and its application on dioxin-containing samples. BMC Bioinform 16: S13. https://doi.org/10.1186/1471-2105-16-S18-S13
|





Figures(6) / Tables(2)

M Thirumala, E Sai Krishna, P Sindhu Priya, S Vishnuvardhan Reddy. Characterization of a novel Fluoride resistant bacterial isolate and its capability of Fluoride bioremediation[J]. AIMS Microbiology, 2022, 8(4): 470-483. doi: 10.3934/microbiol.2022031







 DownLoad:
DownLoad: