Citation: Longxing Qi, Shoujing Tian, Jing-an Cui, Tianping Wang. Multiple infection leads to backward bifurcation for a schistosomiasis model[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 701-712. doi: 10.3934/mbe.2019033

| [1] | T. Britton, T. House, A.L. Lloyd, D. Mollison, S. Riley and P. Trapma, Five challenges for stochastic epidemic models involving global transmission, Epidemics, 10 (2015), 54–57. |
| [2] | Department of disease control of the ministry of health of the people's republic of China, Handbook of schistosomiasis control, Shanghai Science and Technology Press, 2000. (In Chinese) |
| [3] | O. Diekmann, J.A.P. Heesterbeek and J.A.J. Metz, On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations, J. Math. Biol.,28 (1990), 365–382. |
| [4] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [5] | Z.L. Feng, C.C. Li and F.A. Milner, Schistosomiasis models with density dependence and age of infection in snail dynamics, Math. Biosci., 177 (2002), 271–286. |
| [6] | Z.L. Feng, A. Eppert, F.A. Milner and D.J. Minchella, Estimation of parameters governing the transmission dynamics of schistosomes, Appl. Math. Lett., 17 (2004), 1105–1112. |
| [7] | Z.L. Feng, C.C. Li and F.A. Milner, Schistosomiasis models with two migrating human groups, Math. Comput. Model., 41 (2005), 1213–1230. |
| [8] | S.J. Gao, Y.Y. He and Y.J. Liu, Field transmission intensity of Schistosoma japonicum measured by basic reproduction ratio from modified Barbours model, Paras. Vector., 6 (2013), 1–10. |
| [9] | J.H. Ge, S.Q. Zhang and T.P.Wang, Efects of flood On the prevalence of schistosomiasis in Anhui province in 1998, J. Trop. Dis. Parasitol., 2 (2004), 131–134. (In Chinese) |
| [10] | J.C. Kamgang and G. Sallet, Computation of threshold conditions for epidemiological models and global stability of the disease-free equilibrium(DFE), Math. Biosci., 213 (2008), 1–12. |
| [11] | T. Leenstra, H.M. Coutinho, L.P. Acosta, G.C. Langdonet, L. Su, R.M. Olveda, S.T. McGarvey, J.D. Kurtis and J.F. Friedman, Schistosoma japonicum reinfection after Praziquantel treatment causes anemia associated with inflammation, Infect. Immun., 74 (2006), 6398–6407. |
| [12] | S.B. Mao, Biology of schistosome and control of schistosomiasis, People's Health Press, 1990. (In Chinese) |
| [13] | L.X. Qi, J.A. Cui, Y. Gao and H.P. Zhu, Modeling the schistosomiasis on the islets in Nanjing, Int. J. Biomath., 5 (2012), 1–17. |
| [14] | L.X. Qi and J.A. Cui, Mathematical Model of Schistosomiasis under Flood in Anhui Province, Abstr. Appl. Anal., 2014 (2014), 1–7. |
| [15] | S. Riley, H. Carabin, P. Blisle, L. Josephet, V. Tallo, E. Balolong, A.L.Willingham, T.J. Fernandez, R.O. Gonzales, R. Olveda and S.T. McGarvey, Multi-host transmission dynamics of schistosoma japonicum in Samar province, the Philippines, PLoS Med., 5 (2008), 70–78. |
| [16] | WHO, Schistosomiasis, http://www.who.int/mediacentre/factsheets/fs115/en/ |
| [17] | H.J. Xu, X.L. Fu, J.X. Chen, C.M. Xia and J.B. Qiu, Immunopathological observations on the infection of Schistosoma japonicum induced by di erent manners, Chin. J. Zoonoses, 22 (2006), 647–650. |
| [18] | S.Q. Zhang, F.H. Gao, G.H. Zhang, H. Wang and T.P. Wang, Trend analysis of schistosomiasis endemic situation in Anhui Province from 2004 to 2014, Chin. J. Schisto. Control, 27 (2015), 235–240. |
| [19] | G.M. Zhao and J.X. Liu, Study the disease index of schistosoma japonicum infection in the lake region, Chin. J. Schisto. Control, 13 (2001), 173–175. (In Chinese) |
| [20] | L. Zou and S.G. Ruan, Schistosomiasis transmission and control in China, Acta Trop., 143 (2015), 51–57. |
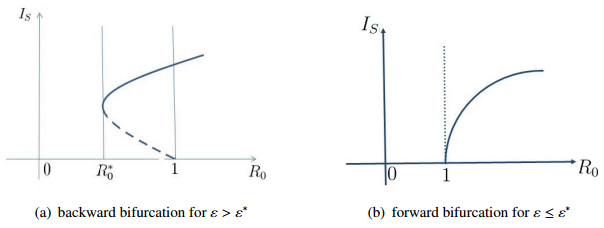
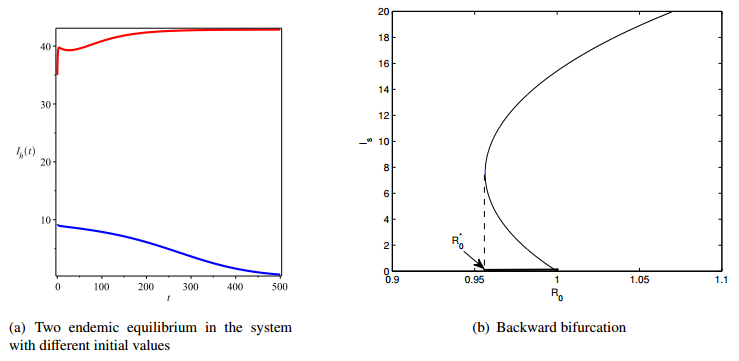
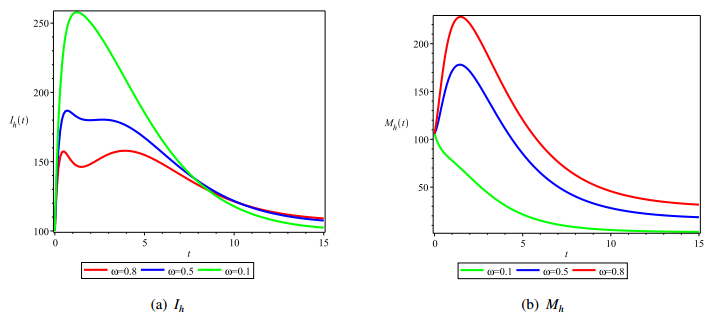
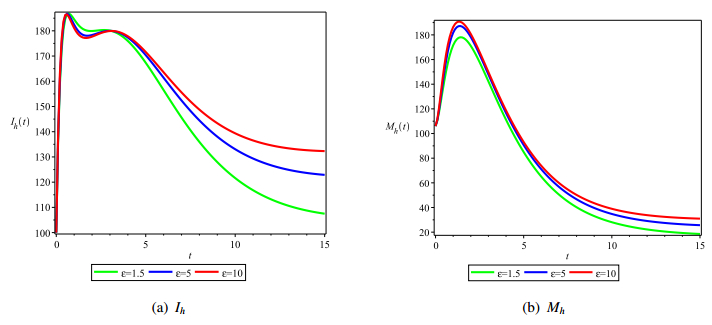
Figures(4) / Tables(1)

Longxing Qi, Shoujing Tian, Jing-an Cui, Tianping Wang. Multiple infection leads to backward bifurcation for a schistosomiasis model[J]. Mathematical Biosciences and Engineering, 2019, 16(2): 701-712. doi: 10.3934/mbe.2019033







 DownLoad:
DownLoad: