Citation: Notice Ringa, Chris T. Bauch. Spatially-implicit modelling of disease-behaviour interactions in the context of non-pharmaceutical interventions[J]. Mathematical Biosciences and Engineering, 2018, 15(2): 461-483. doi: 10.3934/mbe.2018021

| [1] | [ K. A. Alexander,J. W. McNutt, Human behavior influences infectious disease emergence at the human-animal interface, Frontiers in Ecology and the Environment, 8 (2010): 522-526. |
| [2] | [ M. C. Auld, Estimating behavioral response to the AIDS epidemic, Contributions to Economic Analysis and Policy 5 (2006), Art. 12. |
| [3] | [ N. Bacaer, Approximation of the basic reproduction number for vector-borne disease with periodic vector population, Bulleting of Mathematical Biology, 69 (2007): 1067-1091. |
| [4] | [ C. T. Bauch, A versatile ODE approximation to a network model for the spread of sexually transmitted diseases, Journal of Mathematical Biology, 45 (2002): 375-395. |
| [5] | [ C. T. Bauch, The spread of infectious diseases in spatially structured populations: An invasory pair approximation, Mathematical Biosciences, 198 (2005): 217-237. |
| [6] | [ C. T. Bauch,A. d'Onofrio,P. Manfredi, Behavioral epidemiology of infectious diseases: An overview, Modeling the interplay between human behavior and the spread of infectious diseases, Springer-Verlag, null (2013): 1-19. |
| [7] | [ C. T. Bauch,A. P. Galvani, Using network models to approximate spatial point-process models, Mathematical Biosciences, 184 (2003): 101-114. |
| [8] | [ C. T. Bauch,D. A. Rand, A moment closure model for sexually transmitted disease transmission through a concurrent partnership network, The Royal Society, 267 (2000): 2019-2027. |
| [9] | [ J. Benoit,A. Nunes,M. Telo da Gama, Pair approximation models for disease spread, The European Physical Journal B, 50 (2006): 177-181. |
| [10] | [ K. Dietz, The estimation of the basic reproduction number for infectious diseases, Statistical Methods in Medical Research, 2 (1993): 23-41. |
| [11] | [ S. P. Ellner, Pair approximation for lattice models with multiple interaction scales, Journal of Theoretical Biology, 210 (2001): 435-447. |
| [12] | [ E. P. Fenichel,C. Castillo-Chavez,M. G. Geddia,G. Chowell,P. A. Gonzalez Parra,G. J. Hickling,G. Holloway,R. Horan,B. Morin,C. Perrings,M. Springborn,L. Velazquez,C. Villalobos, Adaptive human behavior in epidemiological models, Proceedings of the National Academy of Sciences, 108 (2011): 6306-6311. |
| [13] | [ N. M. Ferguson,C. A. Donnelly,R. M. Anderson, The foot and mouth epidemic in Great Britain: pattern of spread and impact of interventions, Science, 292 (2001): 1155-1160. |
| [14] | [ M. J. Ferrari,S. Bansal,L. A. Meyers,O. N. Bjϕrnstad, Network frailty and the geometry of head immunity, Proceedings of the Royal Society B, 273 (2006): 2743-2748. |
| [15] | [ S. Funk,E. Gilad,C. Watkins,V. A. A. Jansen, The spread of awareness and its impact on epidemic outbreaks, Proceedings of the National Academy of Sciences, 106 (2009): 6872-6877. |
| [16] | [ R. J. Glass,L. M. Glass,W. E. Beyeler,H. J. Min, Targeted social distancing designs for pandemic influenza, Emerging Infectious Diseases, 12 (2016): 1671-1681. |
| [17] | [ D. Hiebeler, Moment equations and dynamics of a household SIS epidemiological model, Bulletin of Mathematical Biology, 68 (2006): 1315-1333. |
| [18] | [ M. J. Keeling, The effects of local spatial structure on epidemiological invasions, Proceedings of The Royal Society of London B, 266 (1999): 859-867. |
| [19] | [ M.J. Keeling,D. A. Rand,A. J. Morris, Correlation models for childhood epidemics, Proceedings of The Royal Society of London B, 264 (1997): 1149-1156. |
| [20] | [ J. Li,D. Blakeley,R. J. Smith, The failure of $\mathbb{R}_0$, Computational and Mathematical Methods in Medicine, 12 (2011): 1-17. |
| [21] | [ C. N. L. Macpherson, Human behavior and the epidemiology of parasitic zoonoses, International Journal for Parasitology, 35 (2005): 1319-1331. |
| [22] | [ S. Maharaj,A. Kleczkowski, Controlling epidemic spread by social distancing: Do it well or not at all, BMC Public Health, 12 (2012): p679. |
| [23] | [ L. Mao,Y. Yang, Coupling infectious diseases, human preventive behavior, and networks--a conceptual framework for epidemic modeling, Social Science Medicine, 74 (2012): 167-175. |
| [24] | [ J. P. McGowan,S. S. Shah,C. E. Ganea,S. Blum,J. A. Ernst,K. L. Irwin,N. Olivo,P. J. Weidle, Risk behavior for transmission of Human Immunodeficiency Virus (HIV) among HIV-seropositive individuals in an urban setting, Clinical Infectious Diseases, 38 (2004): 122-127. |
| [25] | [ T. Modie-Moroka, Intimate partner violence and sexually risky behavior in Botswana: Implications for HIV prevention, Health Care for Women International, 30 (2009): 230-231. |
| [26] | [ S. S. Morse, Factors in the emergence of infectious diseases, Factors in the Emergence of Infectious Diseases, null (2001): 8-26. |
| [27] | [ S. Mushayabasa,C. P. Bhunu,M. Dhlamini, Impact of vaccination and culling on controlling foot and mouth disease: A mathematical modeling approach, World of Journal Vaccines, 1 (2011): 156-161. |
| [28] | [ S. O. Oyeyemi,E. Gabarron,R. Wynn, Ebola, Twitter, and misinformation: A dangerous combination?, British Medical Journal, 349 (2014): g6178. |
| [29] | [ P. E. Parham,N. M. Ferguson, Space and contact networks: Capturing of the locality of disease transmission, Journal of Royal Society, 3 (2005): 483-493. |
| [30] | [ P. E. Parham,B. K. Singh,N. M. Ferguson, Analytical approximation of spatial epidemic models of foot and mouth disease, Theoretical Population Biology, 72 (2008): 349-368. |
| [31] | [ F. M. Pillemer,R. J. Blendon,A. M. Zaslavsky,B. Y. Lee, Predicting support for non-pharmaceutical interventions during infectious outbreaks: A four region analysis, Disasters, 39 (2014): 125-145. |
| [32] | [ D. A. Rand, Correlation equations and pair approximations for spatial ecologies, CWI Quarterly, 12 (1999): 329-368. |
| [33] | [ C. T. Reluga, Game theory of social distancing in response to an epidemic Plos Computational Biology 6 (2010), e1000793, 9pp. |
| [34] | [ N. Ringa,C. T. Bauch, Dynamics and control of foot and mouth disease in endemic countries: A pair approximation model, Journal of Theoretical Biology, 357 (2014): 150-159. |
| [35] | [ A. Rizzo,M. Frasca,M. Porfiri, Effect of individual behavior on epidemic spreading in activity-driven networks, Physical Review E, 90 (2014): 042801. |
| [36] | [ M. Salathe,S. Bonhoeffer, The effect of opinion clustering on disease outbreaks, Journal of The Royal Society Interface, 5 (2008): 1505-1508. |
| [37] | [ L. B. Shaw,I. B. Schwartz, Fluctuating epidemics on adaptive networks, Physical Review E, 77 (2008): 066101, 10pp. |
| [38] | [ R. L. Stoneburner,D. Low-Beer, Population-level HIV decline and behavioral risk avoidance in Uganda, Science, 304 (2004): 714-718. |
| [39] | [ R. R. Swenson,W. S. Hadley,C. D. Houck,S. K. Dance,L. K. Brown, Who accepts a rapid HIV antibody test? The role of race/ethnicity and HIV risk behavior among community adolescents, Journal of Adolescent Health, 48 (2011): 527-529. |
| [40] | [ J. M. Tchuenche and C. T. Bauch, Dynamics of an infectious disease where media coverage influences transmission ISRN Biomathematics2012 (2012), Article ID 581274, 10pp. |
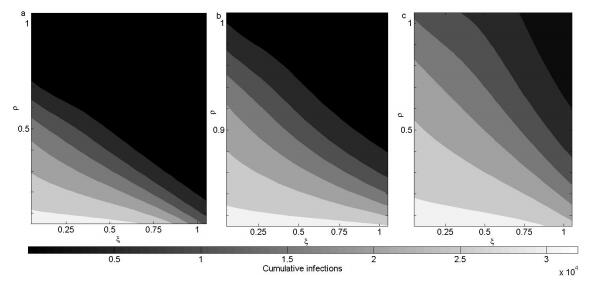
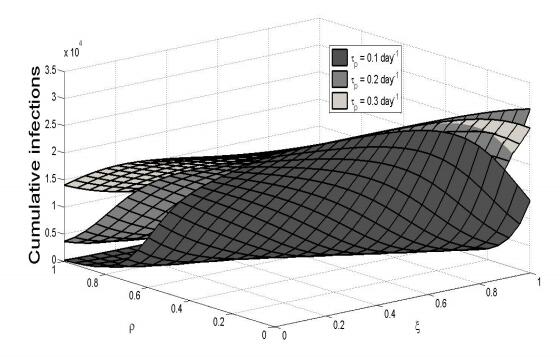
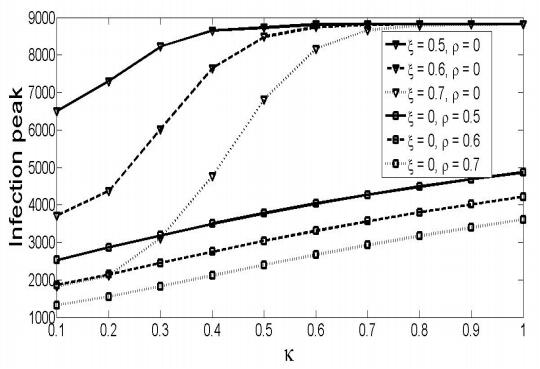
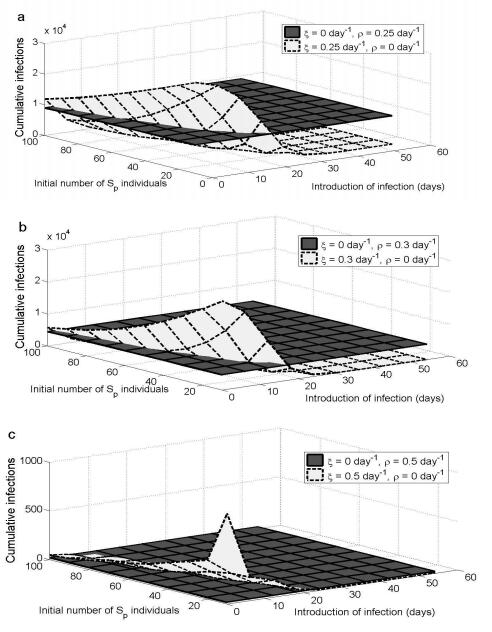
Figures(8) / Tables(1)

Notice Ringa, Chris T. Bauch. Spatially-implicit modelling of disease-behaviour interactions in the context of non-pharmaceutical interventions[J]. Mathematical Biosciences and Engineering, 2018, 15(2): 461-483. doi: 10.3934/mbe.2018021







 DownLoad:
DownLoad: