Citation: Cristina Anton, Jian Deng, Yau Shu Wong, Yile Zhang, Weiping Zhang, Stephan Gabos, Dorothy Yu Huang, Can Jin. Modeling and simulation for toxicity assessment[J]. Mathematical Biosciences and Engineering, 2017, 14(3): 581-606. doi: 10.3934/mbe.2017034

| [1] | [ C. Biernacki,G. Celeux,G. Govaert, Choosing starting values for the EM algorithm for getting the highest likelihood in multivariate gaussian mixture models, Comput. Statist. Data Anal., 41 (2003): 561-575. |
| [2] | [ F. Cannavó, Sensitivity analysis for volcanic source modeling quality assessment and model selection, Computers & Geosciences,, 44 (2012): 52-59. |
| [3] | [ Z. Ghahramani and S. Roweis, Learning nonlinear dynamical systems using an EM algorithm, in Advances in Neural Information Processing Systems (eds. M. Kearns, S. Solla and C. D. A.), MIT Press, 1999,599-605. |
| [4] | [ T. Hallam,C. Clark,G. Jordan, Effects of toxicants on populations: A qualitative approach Ⅱ. First order kinetics, J. Math. Biology, 18 (1983): 25-37. |
| [5] | [ J. He,K. Wang, The survival analysis for a population in a polluted environment, Nonlinear Analysis: Real World Applications, 10 (2009): 1555-1571. |
| [6] | [ B. Huang,J. Xing, Dynamic modeling and prediction of cytotoxicity on microelectronic cell sensor array, The Canadian Journal of Chemical Engineering, 84 (2006): 393-405. |
| [7] | [ Q. Huang,L. Parshotam,H. Wang,C. Bampfylde,M. Lewis, A model for the impact of contaminants on fish population dynamics, Journal of Theoretical Biology, 334 (2013): 71-79. |
| [8] | [ F. Ibrahim,B. Huang,J. Xing,S. Gabos, Early determination of toxicant concentration in water supply using MHE, Water Research, 44 (2010): 3252-3260. |
| [9] | [ A.M. Jarrett,Y. Liu,N. Cogan,M.Y. Hussaini, Global sensitivity analysis used to interpret biological experimental results, Journal of Mathematical Biology, 71 (2015): 151-170. |
| [10] | [ J. Jiao,W. Long,L. Chen, A single stage-structured population model with mature individuals in a polluted environment and pulse input of environmental toxin, Nonlinear Analysis: Real World Applications, 10 (2009): 3073-3081. |
| [11] | [ S. Julier,J. Uhlmann,H. Durrant-White, A new method for the nonlinear transformation of means and covariances in filters and estimators, IEEE Trans. Aut. Control, 45 (2000): 477-482. |
| [12] | [ S. Julier, J. Uhlmann and H. Durrant-Whyte, A new approach for filtering nonlinear systems, in American Control Conference, Seattle, Washington, 1995,1628–1632. |
| [13] | [ A. Kiparissides,S. Kucherenko,A. Mantalaris,E.N. Pistikopoulos, Global sensitivity analysis challenges in biological systems modeling, Industrial & Engineering Chemistry Research, 48 (2009): 7168-7180. |
| [14] | [ K. Kothawad,A. Pathan,M. Logad, Evaluation of in vitro anti-cancer activity of fruit lagenaria siceraria against MCF7, HOP62 and DU145 cell line, Int. J. Pharm. & Technol, 4 (2012): 3909-4392. |
| [15] | [ M. Liu,K. Wang, Survival analysis of stochastic single-species population models in polluted environments, Ecological Modeling, 220 (2009): 1347-1357. |
| [16] | [ M. Liu,K. Wang,X. Liu, Long term behaviors of stochastic single-species growth models in a polluted environment, Applied Mathematical Modelling, 35 (2011): 752-762. |
| [17] | [ X. Meng,D. Van Dyk, The EM algorithm -an old folk-song to a fast new tune, J.R. Statist. Soc.B, 59 (1997): 511-567. |
| [18] | [ R. Neal and G. Hinton, A view of the EM algorithm that justifies incremental, sparse, an other variants, in Learning in Graphical Models (ed. M. Jordan), 89 (1998), 355-368. |
| [19] | [ T. Pan,B. Huang,W. Zhang,S. Gabos,D. Huang,V. Devendran, Cytotoxicity assessment based on the AUC50 using multi-concentration time-dependent cellular response curves, Anal. Chim. Acta, 764 (2013): 44-52. |
| [20] | [ T. Pan,S. Khare,F. Ackah,B. Huang,W. Zhang,S. Gabos,C. Jin,M. Stampfl, In vitro cytotoxicity assessment based on KC50 with real-time cell analyzer (RTCA) assay, Comp. Biol. Chem., 47 (2013): 113-120. |
| [21] | [ L. Perko, Differential Equations and Dynamical Systems, Springer, New York, 2001. |
| [22] | [ R. Shumway,D. Stoffer, An approach to time series smoothing and forecasting using the EM algorithm, J. Time Ser. Anal., 3 (1982): 253-264. |
| [23] | [ I.M. Sobol, Global sensitivity indices for nonlinear mathematical models and their Monte Carlo estimates, Mathematics and Computers in Simulation, 55 (2001): 271-280. |
| [24] | [ H. Thieme, Mathematics in Population Biology, Princeton Series in theoretical and Computational Biology., 2003 |
| [25] | [ E. A. Wan, R. Van der Merwe and A. T. Nelson, Dual estimation and the unscented transformation, in Advances in Neural Information Processing Systems (ed. M. I. J. et al.), MIT Press, 2000. |
| [26] | [ C. Wu, On the convergence properties of the EM algorithm, The Annals of Statistics, 11 (1983): 95-103. |
| [27] | [ Z. Xi,S. Khare,A. Cheung,B. Huang,T. Pan,W. Zhang,F. Ibrahim,C. Jin,S. Gabos, Mode of action classification of chemicals using multi-concentration time-dependent cellular response profiles, Comp. Biol. Chem., 49 (2014): 23-35. |
| [28] | [ J. Xing,L. Zhu,S. Gabos,L. Xie, Microelectronic cell sensor assay for detection of cytotoxicity and prediction of acute toxicity, Toxicology in Vitro, 20 (2006): 995-1004. |
| [29] | [ M. Zhang,D. Aguilera,C. Das,H. Vasquez,P. Zage,V. Gopalakrishnan,J. Wolff, Measuring cytotoxicity: A new perspective on LC50, Anticancer Res., 27 (2007): 35-38. |
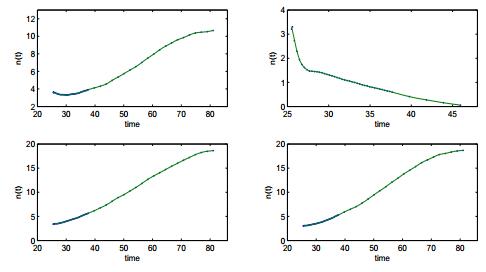
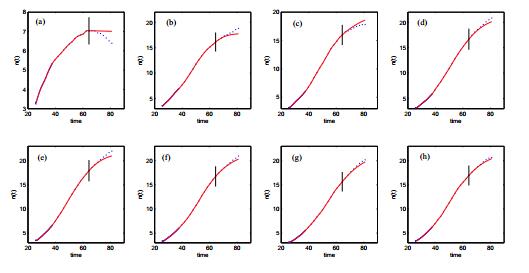
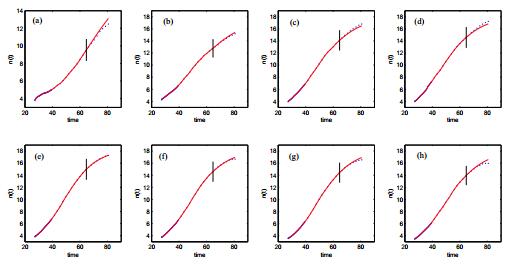
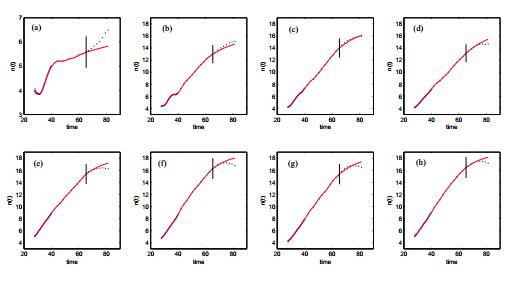
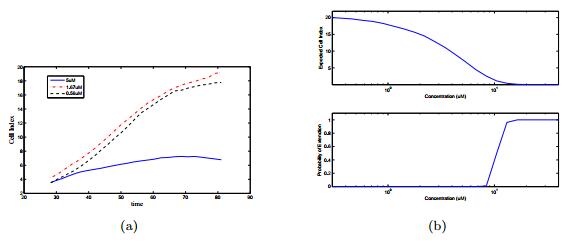
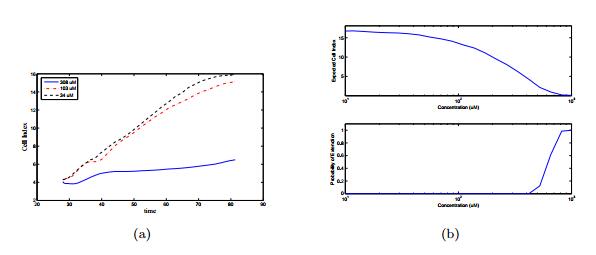
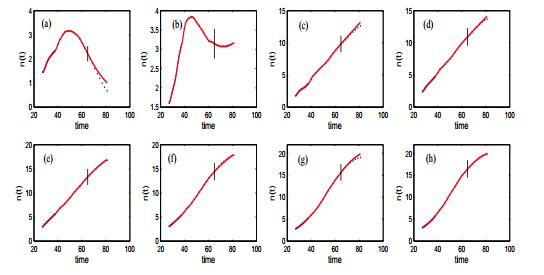
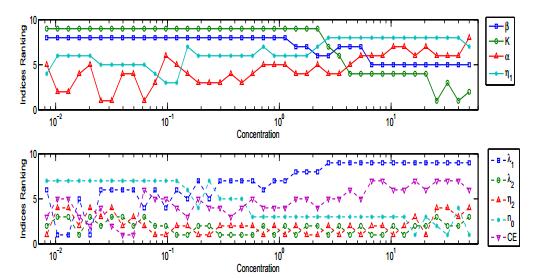
Figures(14) / Tables(3)

Cristina Anton, Jian Deng, Yau Shu Wong, Yile Zhang, Weiping Zhang, Stephan Gabos, Dorothy Yu Huang, Can Jin. Modeling and simulation for toxicity assessment[J]. Mathematical Biosciences and Engineering, 2017, 14(3): 581-606. doi: 10.3934/mbe.2017034







 DownLoad:
DownLoad: