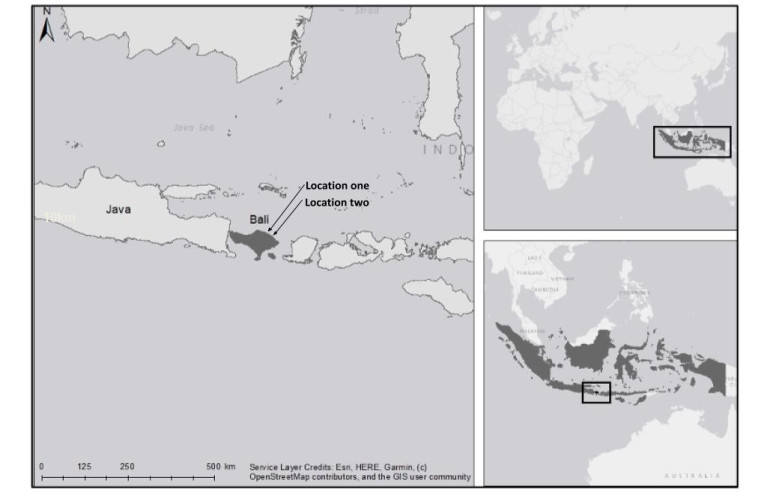
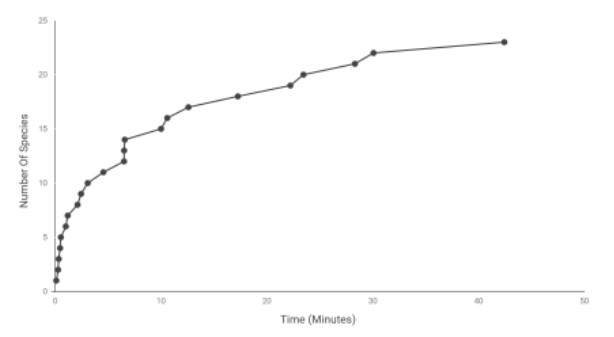
Coral reefs face worldwide decline from threats such as climate change, destructive fishing practices, overfishing and pollution. Artificial reefs have shown potential as a method to mitigate localised habitat loss and biodiversity decline on degraded coral reefs. The health of coral reefs in Indonesia and their associated faunal populations have displayed a downward trend in recent decades, and community-managed non-government organisations have started using artificial reefs to restore local degraded reef habitats. In this study, we demonstrate how locally-managed NGOs and communities in north Bali, Indonesia have implemented artificial reef projects, and assess the associated benefits to biodiversity. Using Remote Underwater Video (RUV) over a 3 month period in north Bali, fish assemblages on two artificial reefs of different ages (new and mature) were compared to two nearby natural habitats: degraded sand flats and relatively healthy coral reefs. When compared with a nearby degraded sand habitat, both artificial reefs displayed a significantly higher number of species, which for the mature artificial reef was not statistically different to a nearby coral reef. Community structure was also compared, again showing similarity between artificial reefs and natural coral reefs, but differing in a few species, including specific damselfish and wrasse. This study is one of few which highlight the potential of artificial reef habitat enhancement in Indonesia, and suggests that these structures can provide ecologically equivalent mobile faunal communities to a natural reef on a localised scale. As such, well designed projects may be able to provide some local ecosystem services lost from degraded coral reefs, and become an important focus for coastal communities.
Citation: Zach Boakes, Alice E Hall, Georgia CA Jones, Rahmadi Prasetijo, Richard Stafford, Yunaldi Yahya. Artificial coral reefs as a localised approach to increase fish biodiversity and abundance along the North Bali coastline[J]. AIMS Geosciences, 2022, 8(2): 303-325. doi: 10.3934/geosci.2022018
Coral reefs face worldwide decline from threats such as climate change, destructive fishing practices, overfishing and pollution. Artificial reefs have shown potential as a method to mitigate localised habitat loss and biodiversity decline on degraded coral reefs. The health of coral reefs in Indonesia and their associated faunal populations have displayed a downward trend in recent decades, and community-managed non-government organisations have started using artificial reefs to restore local degraded reef habitats. In this study, we demonstrate how locally-managed NGOs and communities in north Bali, Indonesia have implemented artificial reef projects, and assess the associated benefits to biodiversity. Using Remote Underwater Video (RUV) over a 3 month period in north Bali, fish assemblages on two artificial reefs of different ages (new and mature) were compared to two nearby natural habitats: degraded sand flats and relatively healthy coral reefs. When compared with a nearby degraded sand habitat, both artificial reefs displayed a significantly higher number of species, which for the mature artificial reef was not statistically different to a nearby coral reef. Community structure was also compared, again showing similarity between artificial reefs and natural coral reefs, but differing in a few species, including specific damselfish and wrasse. This study is one of few which highlight the potential of artificial reef habitat enhancement in Indonesia, and suggests that these structures can provide ecologically equivalent mobile faunal communities to a natural reef on a localised scale. As such, well designed projects may be able to provide some local ecosystem services lost from degraded coral reefs, and become an important focus for coastal communities.

| [1] | Ormond RFG, Roberts CM (1997) The biodiversity of coral reef fishes, Marine Biodiversity Patterns and Process, Cambridge University Press, 460. https://doi.org/10.1017/cbo9780511752360.011 |
| [2] | Spalding M, Spalding MD, Ravilious C, et al. (2001) World atlas of coral reefs, University of California Press. |
| [3] |
Pandolfi JM, Connolly SR, Marshall DJ, et al. (2011) Projecting coral reef futures under global warming and ocean acidification. Science 333: 418-422. https://doi.org/10.1126/science.1204794 doi: 10.1126/science.1204794

|
| [4] |
Hughes TP, Kerry JT, Baird AH, et al. (2018) Global warming transforms coral reef assemblages. Nature 556: 492-496. https://doi.org/10.1038/s41586-018-0041-2 doi: 10.1038/s41586-018-0041-2
|
| [5] |
Richardson LE, Graham NAJ, Pratchett MS, et al. (2018) Mass coral bleaching causes biotic homogenization of reef fish assemblages. Glob Chang Biol 24: 3117-3129. https://doi.org/10.1111/gcb.14119 doi: 10.1111/gcb.14119
|
| [6] |
Bell JD, Ratner BD, Stobutzki I, et al. (2006) Addressing the coral reef crisis in developing countries. Ocean Coastal Manage 49: 976-985. https://doi.org/10.1016/j.ocecoaman.2005.12.001 doi: 10.1016/j.ocecoaman.2005.12.001
|
| [7] |
Moberg F, Folke C (1999) Ecological goods and services of coral reef ecosystems. Ecol Econ 29: 215-233. http://dx.doi.org/10.1016/S0921-8009(99)00009-9 doi: 10.1016/S0921-8009(99)00009-9
|
| [8] | Principe P, Bradley P, Yee S, et al. (2012) Quantifying coral reef ecosystem services. US Environmental Protection Agency, NC. Available from: https://cfpub.epa.gov/si/si_public_record_report.cfm?Lab=NERL&dirEntryId=239984 |
| [9] | Heron SF, Eakin CM, Douvere F, et al. (2017) Impacts of climate change on World Heritage coral reefs: a first global scientific assessment. Available from: https://repository.library.noaa.gov/view/noaa/16386 |
| [10] | Pörtner HO, Karl DM, Boyd PW, et al. (2014) Ocean systems, Climate change 2014: impacts, adaptation, and vulnerability. Part A: global and sectoral aspects. contribution of working group II to the fifth assessment report of the intergovernmental panel on climate change, Cambridge University Press, 411-484. |
| [11] |
Vivier B, Dauvin JC, Navon M, et al. (2021) Marine artificial reefs, a meta-analysis of their design, objectives and effectiveness. Glob Ecol Conserv 27: e01538. https://doi.org/10.1016/j.gecco.2021.e01538 doi: 10.1016/j.gecco.2021.e01538
|
| [12] |
Paxton AB, Shertzer KW, Bacheler NM, et al. (2020) Meta-analysis reveals artificial reefs can be effective tools for fish community enhancement but are not one-size-fits-all. Front Mar Sci 7: 282. https://doi.org/10.3389/fmars.2020.00282 doi: 10.3389/fmars.2020.00282
|
| [13] |
Messmer V, Jones GP, Munday PL, et al. (2011) Habitat biodiversity as a determinant of fish community structure on coral reefs. Ecology 92: 2285-2298. https://doi.org/10.1890/11-0037.1 doi: 10.1890/11-0037.1
|
| [14] |
Komyakova V, Munday PL, Jones GP (2013) Relative importance of coral cover, habitat complexity and diversity in determining the structure of reef fish communities. PLoS One 8: e83178. https://doi.org/10.1371/journal.pone.0083178 doi: 10.1371/journal.pone.0083178
|
| [15] |
Becker A, Taylor MD, Lowry MB (2017) Monitoring of reef associated and pelagic fish communities on Australia's first purpose built offshore artificial reef. ICES J Mar Sci 74: 277-285. https://doi.org/10.1093/icesjms/fsw133 doi: 10.1093/icesjms/fsw133
|
| [16] |
Israel D, Gallo C, Angel DL (2017) Benthic artificial reefs as a means to reduce the environmental effects of cod mariculture in Skutulsfjörð ur, Iceland. Mar Biodiv 47: 405-411. https://doi.org/10.1007/s12526-016-0490-5 doi: 10.1007/s12526-016-0490-5
|
| [17] |
Baine M (2001) Artificial reefs: a review of their design, application, management and performance. Ocean Coast Manage 44: 241-259. https://doi.org/10.1016/S0964-5691(01)00048-5 doi: 10.1016/S0964-5691(01)00048-5
|
| [18] | Bohnsack JA, Sutherland DL (1985) Artificial reef research: a review with recommendations for future priorities. Bull Mar Sci 37: 11-39. |
| [19] |
Keller K, Steffe AS, Lowry MB, et al. (2017) Estimating the recreational harvest of fish from a nearshore designed artificial reef using a pragmatic approach. Fish Res 187: 158-167. https://doi.org/10.1016/j.fishres.2016.11.022 doi: 10.1016/j.fishres.2016.11.022
|
| [20] |
Kirkbride-Smith AE, Wheeler PM, Johnson ML (2016) Artificial reefs and marine protected areas: a study in willingness to pay to access Folkestone Marine Reserve, Barbados, West Indies. PeerJ 4: e2175. https://doi.org/10.7717/peerj.2175 doi: 10.7717/peerj.2175
|
| [21] | Bideci C, Cater C (2019) In search of underwater atmosphere: A new diving world on artificial reefs, Atmospheric turn in culture and tourism: Place, design and process impacts on customer behaviour, marketing and branding, Emerald Publishing Limited. |
| [22] | Relini M, Torchia G, Relini G (1994) Seasonal variation of fish assemblages in the Loano artificial reef (Ligurian Sea Northwestern-Mediterranean). Bull Mar Sci 55: 401-417. |
| [23] | Oh TG, Otake S, Lee MO (2011) Estimating the effective wake region (current shadow) of artificial reefs. Artificial Reefs in Fisheries Management, Eds., 279-295. https://doi.org/10.1201/b10910-21 |
| [24] |
Herbert RJH, Collins K, Mallinson J, et al. (2017) Epibenthic and mobile species colonisation of a geotextile artificial surf reef on the south coast of England. PLoS One 12: e0184100. https://doi.org/10.1371/journal.pone.0184100 doi: 10.1371/journal.pone.0184100
|
| [25] |
Perkol-Finkel S, Shashar N, Benayahu Y (2006) Can artificial reefs mimic natural reef communities? The roles of structural features and age. Mar Environ Res 61: 121-135. https://doi.org/10.1016/j.marenvres.2005.08.001 doi: 10.1016/j.marenvres.2005.08.001
|
| [26] | Marinaro JY (1995) Artificial reefs in the French Mediterranean: A critical assessment of previous experiments and a proposition in favour of a new reef-planning policy. Biol Mar Mediterr 2: 65-76. |
| [27] |
Lemoine HR, Paxton AB, Anisfeld SC, et al. (2019) Selecting the optimal artificial reefs to achieve fish habitat enhancement goals. Biol Conserv 238: 108200. https://doi.org/10.1016/j.biocon.2019.108200 doi: 10.1016/j.biocon.2019.108200
|
| [28] |
Komyakova V, Chamberlain D, Jones GP, et al. (2019) Assessing the performance of artificial reefs as substitute habitat for temperate reef fishes: Implications for reef design and placement. Sci Total Environ 668: 139-152. https://doi.org/10.1016/j.scitotenv.2019.02.357 doi: 10.1016/j.scitotenv.2019.02.357
|
| [29] | Bailey-Brock JH (1989) Fouling community development on an artificial reef in Hawaiian waters. Bull Mar Sci 44: 580-591. |
| [30] |
Pickering H, Whitmarsh D (1997) Artificial reefs and fisheries exploitation: a review of the 'attraction versus production' debate, the influence of design and its significance for policy. Fish Res 31: 39-59. https://doi.org/10.1016/S0165-7836(97)00019-2 doi: 10.1016/S0165-7836(97)00019-2
|
| [31] |
Arney RN, Froehlich CYM, Kline RJ (2017) Recruitment patterns of juvenile fish at an artificial reef area in the Gulf of Mexico. Mar Coast Fish 9: 79-92. https://doi.org/10.1080/19425120.2016.1265031 doi: 10.1080/19425120.2016.1265031
|
| [32] |
Addis DT, Patterson Iii WF, Dance MA (2016) The potential for unreported artificial reefs to serve as refuges from fishing mortality for reef fishes. N Am J Fish Manage 36: 131-139. https://doi.org/10.1080/02755947.2015.1084406 doi: 10.1080/02755947.2015.1084406
|
| [33] |
Carr MH, Hixon MA (1997) Artificial reefs: the importance of comparisons with natural reefs. Fisheries 22: 28-33. https://doi.org/10.1577/1548-8446(1997)022<0028:ARTIOC>2.0.CO;2 doi: 10.1577/1548-8446(1997)022<0028:ARTIOC>2.0.CO;2
|
| [34] |
Folpp H, Lowry M, Gregson M, et al. (2013) Fish assemblages on estuarine artificial reefs: natural rocky-reef mimics or discrete assemblages? PLoS One 8: e63505. https://doi.org/10.1371/journal.pone.0063505 doi: 10.1371/journal.pone.0063505
|
| [35] |
Brickhill MJ, Lee SY, Connolly RM (2005) Fishes associated with artificial reefs: attributing changes to attraction or production using novel approaches. J Fish Biol 67: 53-71. https://doi.org/10.1111/j.0022-1112.2005.00915.x doi: 10.1111/j.0022-1112.2005.00915.x
|
| [36] |
Smith JA, Lowry MB, Suthers IM (2015) Fish attraction to artificial reefs not always harmful: a simulation study. Ecol Evol 5: 4590-4602. https://doi.org/10.1002/ece3.1730 doi: 10.1002/ece3.1730
|
| [37] |
Roa-Ureta RH, Santos MN, Leitã o F (2019) Modelling long-term fisheries data to resolve the attraction versus production dilemma of artificial reefs. Ecol Modell 407: 108727. https://doi.org/10.1016/j.ecolmodel.2019.108727 doi: 10.1016/j.ecolmodel.2019.108727
|
| [38] | Pears RJ, Williams DM (2005) Potential effects of artificial reefs on the Great Barrier Reef: background paper. CRC Reef Research Centre Townsville, Australia. Available from: http://www.reef.crc.org.au/ |
| [39] |
Heery EC, Bishop MJ, Critchley LP, et al. (2017) Identifying the consequences of ocean sprawl for sedimentary habitats. J Exp Mar Biol Ecol 492: 31-48. https://doi.org/10.1016/j.jembe.2017.01.020 doi: 10.1016/j.jembe.2017.01.020
|
| [40] |
Blount C, Komyakova V, Barnes L, et al. (2021) Using ecological evidence to refine approaches to deploying offshore artificial reefs for recreational fisheries. Bull Mar Sci 97: 665-698. https://doi.org/10.5343/bms.2020.0059 doi: 10.5343/bms.2020.0059
|
| [41] |
Sheehy DJ, Vik SF (2010) The role of constructed reefs in non-indigenous species introductions and range expansions. Ecol Eng 36: 1-11. https://doi.org/10.1016/j.ecoleng.2009.09.012 doi: 10.1016/j.ecoleng.2009.09.012
|
| [42] |
Sutton SG, Bushnell SL (2007) Socio-economic aspects of artificial reefs: Considerations for the Great Barrier Reef Marine Park. Ocean Coast Manage 50: 829-846. https://doi.org/10.1016/j.ocecoaman.2007.01.003 doi: 10.1016/j.ocecoaman.2007.01.003
|
| [43] |
Cullen-Knox C, Haward M, Jabour J, et al. (2017) The social licence to operate and its role in marine governance: insights from Australia. Mar Policy 79: 70-77. https://doi.org/10.1016/j.marpol.2017.02.013 doi: 10.1016/j.marpol.2017.02.013
|
| [44] |
Lima JS, Zalmon IR, Love M (2019) Overview and trends of ecological and socioeconomic research on artificial reefs. Mar Environ Res 145: 81-96. https://doi.org/10.1016/j.marenvres.2019.01.010 doi: 10.1016/j.marenvres.2019.01.010
|
| [45] |
Voyer M, Gladstone W, Goodall H (2015) Obtaining a social licence for MPAs—influences on social acceptability. Mar Policy 51: 260-266. https://doi.org/10.1016/j.marpol.2014.09.004 doi: 10.1016/j.marpol.2014.09.004
|
| [46] |
Bennett NJ, Dearden P (2014) Why local people do not support conservation: Community perceptions of marine protected area livelihood impacts, governance and management in Thailand. Mar policy 44: 107-116. https://doi.org/10.1016/j.marpol.2013.08.017 doi: 10.1016/j.marpol.2013.08.017
|
| [47] | Berkes F (2010) Linkages and multilevel systems for matching governance and ecology: lessons from roving bandits. Bull Mar Sci 86: 235-250. |
| [48] |
Lundquist CJ, Granek EF (2005) Strategies for successful marine conservation: integrating socioeconomic, political, and scientific factors. Conserv Biol 19: 1771-1778. https://doi.org/10.1111/j.1523-1739.2005.00279.x doi: 10.1111/j.1523-1739.2005.00279.x
|
| [49] |
Glaser M, Baitoningsih W, Ferse SCA, et al. (2010) Whose sustainability? Top-down participation and emergent rules in marine protected area management in Indonesia. Mar Policy 34: 1215-1225. https://doi.org/10.1016/j.marpol.2010.04.006 doi: 10.1016/j.marpol.2010.04.006
|
| [50] |
Trialfhianty TI (2017) The role of the community in supporting coral reef restoration in Pemuteran, Bali, Indonesia. J Coast Conserv 21: 873-882. https://doi.org/10.1007/s11852-017-0553-1 doi: 10.1007/s11852-017-0553-1
|
| [51] |
Strain EMA, Alexander KA, Kienker S, et al. (2019) Urban blue: a global analysis of the factors shaping people's perceptions of the marine environment and ecological engineering in harbours. Sci Total Environ 658: 1293-1305. https://doi.org/10.1016/j.scitotenv.2018.12.285 doi: 10.1016/j.scitotenv.2018.12.285
|
| [52] |
Di Lorenzo M, Guidetti P, Di Franco A, et al. (2020) Assessing spillover from marine protected areas and its drivers: A meta‐analytical approach. Fish Fish 21: 906-915. https://doi.org/10.1111/faf.12469 doi: 10.1111/faf.12469
|
| [53] |
Lenihan HS, Gallagher JP, Peters JR, et al. (2021) Evidence that spillover from Marine Protected Areas benefits the spiny lobster (Panulirus interruptus) fishery in southern California. Sci Rep 11: 1-9. https://doi.org/10.1038/s41598-021-82371-5 doi: 10.1038/s41598-021-82371-5
|
| [54] |
King AJ, George A, Buckle DJ, et al. (2018) Efficacy of remote underwater video cameras for monitoring tropical wetland fishes. Hydrobiologia 807: 145-164. https://doi.org/10.1007/s10750-017-3390-1 doi: 10.1007/s10750-017-3390-1
|
| [55] |
Hall AE, Herbert RJH, Stafford R (2021) Temporal and spatial variation in adult and juvenile mobile fauna associated with natural and artificial coastal habitats. Mar Biol 168: 1-13. https://doi.org/10.1007/s00227-021-03823-0 doi: 10.1007/s00227-021-03823-0
|
| [56] | Allen GR, Steene R, Humann P, et al. (2003) Reef fish identification: tropical Pacific, New World Publications Incorporated. |
| [57] |
Whitmarsh SK, Fairweather PG, Huveneers C (2017) What is Big BRUVver up to? Methods and uses of baited underwater video. Rev Fish Biol Fisherise 27: 53-73. https://doi.org/10.1007/s11160-016-9450-1 doi: 10.1007/s11160-016-9450-1
|
| [58] |
Schramm KD, Harvey ES, Goetze JS, et al. (2020) A comparison of stereo-BRUV, diver operated and remote stereo-video transects for assessing reef fish assemblages. J Exp Mar Bio Ecol 524: 151273. https://doi.org/10.1016/j.jembe.2019.151273 doi: 10.1016/j.jembe.2019.151273
|
| [59] |
Bates D, Mächler M, Bolker B, et al. (2014) Fitting linear mixed-effects models using lme4. J Stat Software 67: 1-48. https://doi.org/10.18637/jss.v067.i01 doi: 10.18637/jss.v067.i01
|
| [60] | Howlett SJ, Stafford R, Waller M, et al. (2016) Linking protection with the distribution of grouper and habitat quality in Seychelles. J Mar Biol 2016. https://doi.org/10.1155/2016/7851425 |
| [61] |
Anderson MJ (2001) A new method for non‐parametric multivariate analysis of variance. Austral Ecol 26: 32-46. https://doi.org/10.1111/j.1442-9993.2001.01070.pp.x doi: 10.1111/j.1442-9993.2001.01070.pp.x
|
| [62] | Nakamura M (1985). Evolution of artificial fishing reef concepts in Japan. Bull Mar Sci 37: 271-278. Available from: https://jglobal.jst.go.jp/en/detail?JGLOBAL_ID=201002058198238679 |
| [63] |
Hackradt CW, Félix-Hackradt FC, García-Charton JA (2011) Influence of habitat structure on fish assemblage of an artificial reef in southern Brazil. Mar Environ Res 72: 235-247. https://doi.org/10.1016/j.marenvres.2011.09.006 doi: 10.1016/j.marenvres.2011.09.006
|
| [64] | Kingsford MJ, Leis JM, Shanks A, et al. (2002) Sensory environments, larval abilities and local self-recruitment. Bull Mar Sci 70: 309-340. |
| [65] |
Folpp HR, Schilling HT, Clark GF, et al. (2020) Artificial reefs increase fish abundance in habitat‐limited estuaries. J Appl Ecol 57: 1752-1761. https://doi.org/10.1111/1365-2664.13666 doi: 10.1111/1365-2664.13666
|
| [66] | Doherty PJ (1983) Tropical territorial damselfishses: is density limited by aggression or recruitment? Ecology 64: 176-190. |
| [67] |
Frédérich B, Fabri G, Lepoint G, et al. (2009) Trophic niches of thirteen damselfishes (Pomacentridae) at the Grand Récif of Toliara, Madagascar. Ichthyol Res 56: 10-17. https://doi.org/10.1007/s10228-008-0053-2 doi: 10.1007/s10228-008-0053-2
|
| [68] |
Depczynski M, Bellwood DR (2004) Microhabitat utilisation patterns in cryptobenthic coral reef fish communities. Mar Biol 145: 455-463. https://doi.org/10.1007/s00227-004-1342-6 doi: 10.1007/s00227-004-1342-6
|
| [69] | Wulff JL (1994) Sponge feeding by Caribbean angelfishes, trunkfishes, and filefishes. Sponges Time Space, 265-271. Availble from: https://www.bio.fsu.edu/wulff/Wulff%201994a.pdf |
| [70] |
Seemann J, Yingst A, Stuart-Smith RD, et al. (2018) The importance of sponges and mangroves in supporting fish communities on degraded coral reefs in Caribbean Panama. PeerJ 6: e4455. https://doi.org/10.7717/peerj.4455 doi: 10.7717/peerj.4455
|
| [71] |
Brandl SJ, Rasher DB, Côté IM, et al. (2019) Coral reef ecosystem functioning: eight core processes and the role of biodiversity. Front Ecol Environ 17: 445-454. https://doi.org/10.1002/fee.2088 doi: 10.1002/fee.2088
|
| [72] |
McCauley DJ, Young HS, Dunbar RB, et al. (2012) Assessing the effects of large mobile predators on ecosystem connectivity. Ecol Appl 22: 1711-1717. https://doi.org/10.1890/11-1653.1 doi: 10.1890/11-1653.1
|
| [73] |
Kakizaki T, Kobayashi K, Nakatsubo T, et al. (2015) Spawning behavior of garden eels, Gorgasia preclara and Heteroconger hassi (Heterocongrinae), observed in captivity. Mar Freshw Behav Physiol 48: 359-373. https://doi.org/10.1080/10236244.2015.1064213 doi: 10.1080/10236244.2015.1064213
|
| [74] |
Kessel ST, Hinojosa NA, Wilson H, et al. (2018) Varied response of garden eels to potential predators and other large-bodied organisms. Matters 4: 1-5. https://doi.org/10.19185/matters.201806000024 doi: 10.19185/matters.201806000024
|
| [75] |
Huijbers CM, Nagelkerken I, Layman CA (2015) Fish movement from nursery bays to coral reefs: a matter of size? Hydrobiologia 750: 89-101. https://doi.org/10.1007/s10750-014-2162-4 doi: 10.1007/s10750-014-2162-4
|
| [76] |
Mills KA, Hamer PA, Quinn GP (2017) Artificial reefs create distinct fish assemblages. Mar Ecol Prog Ser 585: 155-173. https://doi.org/10.3354/meps12390 doi: 10.3354/meps12390
|
| [77] | Syam AR, Edrus IN, Hartati ST (2017) Coral fish population changes in the surrounding artificial reefs of the Lebah coastal waters, Karangasem, Bali. Ind Fish Res J 13: 101-116. |




Figures(6) / Tables(4)

Zach Boakes, Alice E Hall, Georgia CA Jones, Rahmadi Prasetijo, Richard Stafford, Yunaldi Yahya. Artificial coral reefs as a localised approach to increase fish biodiversity and abundance along the North Bali coastline[J]. AIMS Geosciences, 2022, 8(2): 303-325. doi: 10.3934/geosci.2022018







 DownLoad:
DownLoad: