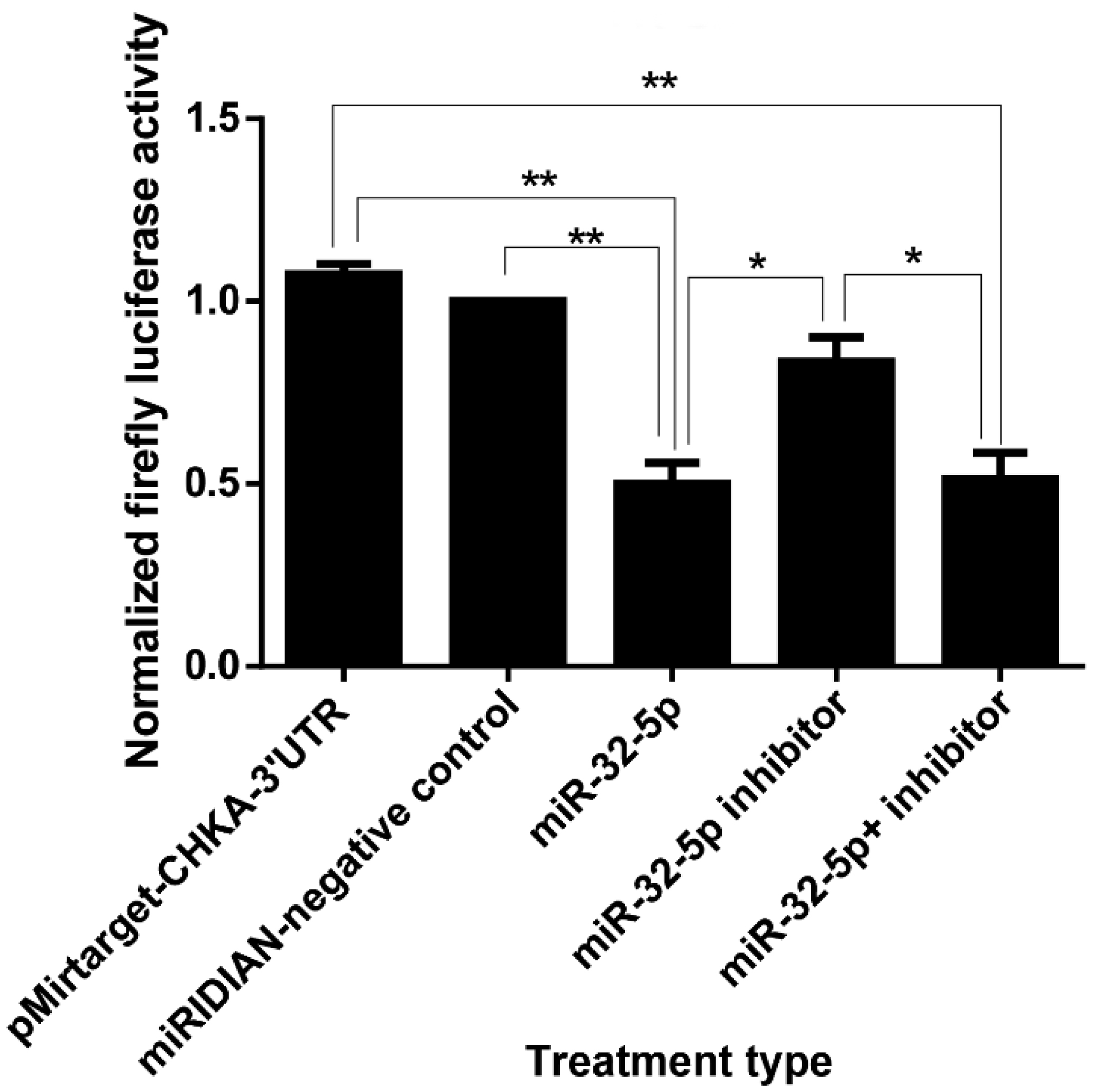
Cervical cancer remains a significant global health challenge, especially in low- and middle-income countries. The elucidation of the molecular pathways underlying the carcinogenesis of cervical cancer is vital to develop targeted therapies. Our study investigates the regulatory effects of miR-32-5p on the choline kinase alpha (chka) gene in HeLa cells, a well-established cervical cancer cell line. Choline kinase alpha is recognized for its role in phosphatidylcholine biosynthesis, which is crucial for cell membrane formation, and is implicated in the oncogenic transformation of cells. Utilizing a combination of in silico prediction, luciferase assays, RT-qPCR, and Western blot analyses, we demonstrated that miR-32-5p directly targets the 3′ untranslated region (3′UTR) of chka mRNA, leading to a significant downregulation of chka expression. Our results demonstrate that miR-32-5p significantly downregulates chka at both the mRNA and protein levels, thus leading to decreased cellular proliferation and increased apoptosis. This was further confirmed by a cell cycle analysis, which showed a notable arrest in the G0/G1 phase. Additionally, scratch assays indicated a reduced migratory capacity in miR-32-5p-transfected cells, suggesting the potential anti-metastatic properties of miR-32-5p. These findings highlight the therapeutic potential of miR-32-5p as a biomarker and a target in cervical cancer treatment strategies. By modulating chka expression, miR-32-5p could serve as a novel approach to curb the progression and spread of cervical cancer, thus offering a promising avenue for future research and clinical applications. This study contributes to the growing understanding of miRNA-mediated gene regulation in cancer biology and underscores the importance of targeted genetic research in the development of cancer therapeutics.
Citation: Sweta Raikundalia, Ling Ling Few, Siti Asma' Hassan, Get Bee Yvonne-Τee, Wei Cun See Too. Choline kinase and miR-32-5p: A crucial interaction promoting apoptosis and delaying wound repair in cervical cancer cells[J]. AIMS Biophysics, 2024, 11(3): 281-295. doi: 10.3934/biophy.2024016
Cervical cancer remains a significant global health challenge, especially in low- and middle-income countries. The elucidation of the molecular pathways underlying the carcinogenesis of cervical cancer is vital to develop targeted therapies. Our study investigates the regulatory effects of miR-32-5p on the choline kinase alpha (chka) gene in HeLa cells, a well-established cervical cancer cell line. Choline kinase alpha is recognized for its role in phosphatidylcholine biosynthesis, which is crucial for cell membrane formation, and is implicated in the oncogenic transformation of cells. Utilizing a combination of in silico prediction, luciferase assays, RT-qPCR, and Western blot analyses, we demonstrated that miR-32-5p directly targets the 3′ untranslated region (3′UTR) of chka mRNA, leading to a significant downregulation of chka expression. Our results demonstrate that miR-32-5p significantly downregulates chka at both the mRNA and protein levels, thus leading to decreased cellular proliferation and increased apoptosis. This was further confirmed by a cell cycle analysis, which showed a notable arrest in the G0/G1 phase. Additionally, scratch assays indicated a reduced migratory capacity in miR-32-5p-transfected cells, suggesting the potential anti-metastatic properties of miR-32-5p. These findings highlight the therapeutic potential of miR-32-5p as a biomarker and a target in cervical cancer treatment strategies. By modulating chka expression, miR-32-5p could serve as a novel approach to curb the progression and spread of cervical cancer, thus offering a promising avenue for future research and clinical applications. This study contributes to the growing understanding of miRNA-mediated gene regulation in cancer biology and underscores the importance of targeted genetic research in the development of cancer therapeutics.

| [1] | World Health Organization Global Health Estimates 2020: Deaths by Cause, Age, sex, by Country and by Region, 2000−2019, 2023. Available from: https://www.who.int/data/gho/data/themes/mortality-and-global-health-estimates/ghe-leading-causes-of-death |
| [2] |
Sung H, Ferlay J, Siegel RL, et al. (2021) Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 71: 209-249. https://doi.org/10.3322/caac.21660 
|
| [3] |
Ramírez de Molina A, Gallego-Ortega D, Sarmentero J, et al. (2005) Choline kinase is a novel oncogene that potentiates RhoA-induced carcinogenesis. Cancer Res 65: 5647-5653. https://doi.org/10.1158/0008-5472.CAN-04-4416
|
| [4] |
Lacal Sanjuán JC, Zimmerman T, Campos Rosa JM (2021) Choline kjinase: An unexpected journey for a precision medicine strategy in human diseases. Pharmaceutics 13: 788. https://doi.org/10.3390/pharmaceutics13060788
|
| [5] |
Rizzo A, Satta A, Garrone G, et al. (2021) Choline kinase alpha impairment overcomes TRAIL resistance in ovarian cancer cells. J Exp Clin Cancer Res 40: 5. https://doi.org/10.1186/s13046-020-01794-6
|
| [6] |
Falcon SC, Hudson CS, Huang Y, et al. (2013) A non-catalytic role of choline kinase alpha is important in promoting cancer cell survival. Oncogenesis 2: e38. https://doi.org/10.1038/oncsis.2013.2
|
| [7] |
Trousil S, Kaliszczak M, Schug Z, et al. (2016) The novel choline kinase inhibitor ICL-CCIC-0019 reprograms cellular metabolism and inhibits cancer cell growth. Oncotarget 7: 37103-37120. https://doi.org/10.18632/oncotarget.9466
|
| [8] |
Sola-Leyva A, López-Cara LC, Ríos-Marco P, et al. (2019) Choline kinase inhibitors EB-3D and EB-3P interferes with lipid homeostasis in HepG2 cells. Sci Rep 9: 5109. https://doi.org/10.1038/s41598-019-40885-z
|
| [9] |
Shah V, Shah J (2020) Recent trends in targeting miRNAs for cancer therapy. J Pharm Pharmacol 72: 1732-1749. https://doi.org/10.1111/jphp.13351
|
| [10] | Lan H, Lu H, Wang X, et al. (2015) MicroRNAs as potential biomarkers in cancer: opportunities and challenges. Biomed Res Int 2015: 125094. https://doi.org/10.1155/2015/125094 |
| [11] |
Arlauckas SP, Popov AV, Delikatny EJ (2016) Choline kinase alpha—Putting the ChoK-hold on tumor metabolism. Prog Lipid Res 63: 28-40. https://doi.org/10.1016/j.plipres.2016.03.005
|
| [12] |
Gruber J, See Too WC, Wong MT, et al. (2012) Balance of human choline kinase isoforms is critical for cell cycle regulation: implications for the development of choline kinase-targeted cancer therapy. FEBS J 279: 1915-1928. https://doi.org/10.1111/j.1742-4658.2012.08573.x
|
| [13] |
Ramírez de Molina A, Gutiérrez R, Ramos MA, et al. (2002) Increased choline kinase activity in human breast carcinomas: clinical evidence for a potential novel antitumor strategy. Oncogene 21: 4317-4322. https://doi.org/10.1038/sj.onc.1205556
|
| [14] |
Clem BF, Clem AL, Yalcin A, et al. (2011) A novel small molecule antagonist of choline kinase-α that simultaneously suppresses MAPK and PI3K/AKT signaling. Oncogene 30: 3370-3380. https://doi.org/10.1038/onc.2011.51
|
| [15] |
Gokhale S, Xie P (2021) ChoK-full of potential: Choline kinase in B cell and T cell malignancies. Pharmaceutics 13: 911. https://doi.org/10.3390/pharmaceutics13060911
|
| [16] |
Crilly KS, Tomono M, Kiss Z (1998) The choline kinase inhibitor hemicholinium-3 can inhibit mitogen-induced DNA synthesis independent of its effect on phosphocholine formation. Arch Biochem Biophys 352: 137-143. https://doi.org/10.1006/abbi.1998.0601
|
| [17] |
Schiaffino-Ortega S, Baglioni E, Mariotto E, et al. (2016) Design, synthesis, crystallization and biological evaluation of new symmetrical biscationic compounds as selective inhibitors of human Choline Kinase α1 (ChoKα1). Sci Rep 6: 23793. https://doi.org/10.1038/srep23793
|
| [18] |
Lin XM, Hu L, Gu J, et al. (2017) Choline kinase α mediates interactions between the epidermal growth factor receptor and mechanistic target of rapamycin complex 2 in hepatocellular carcinoma cells to promote drug resistance and xenograft tumor progression. Gastroenterology 152: 1187-1202. https://doi.org/10.1053/j.gastro.2016.12.033
|
| [19] |
Barthe A, Bodaar K, Issaian A, et al. (2022) Choline kinase a is required for alkylating agent resistance in T-cell acute lymphoblastic leukemia. Blood 140: 2258. https://doi.org/10.1182/blood-2022-162508
|
| [20] |
Qin SY, Li B, Chen M, et al. (2022) MiR-32-5p promoted epithelial-to-mesenchymal transition of oral squamous cell carcinoma cells via regulating the KLF2/CXCR4 pathway. Kaohsiung J Med Sci 38: 120-128. https://doi.org/10.1002/kjm2.12450
|
| [21] |
Zeng S, Liu S, Feng J, et al. (2020) MicroRNA-32 promotes ovarian cancer cell proliferation and motility by targeting SMG1. Oncol Lett 20: 733-741. https://doi.org/10.3892/ol.2020.11624
|
| [22] |
Zhang JX, Yang W, Wu JZ, et al. (2021) MicroRNA-32-5p inhibits epithelial-mesenchymal transition and metastasis in lung adenocarcinoma by targeting SMAD family 3. J Cancer 12: 2258-2267. https://doi.org/10.7150/jca.48387
|
| [23] | Liu YJ, Zhou HG, Chen LH, et al. (2019) MiR-32-5p regulates the proliferation and metastasis of cervical cancer cells by targeting HOXB8. Eur Rev Med Pharmacol Sci 23: 87-95. https://doi.org/10.26355/eurrev_201901_16752 |




Figures(5)

Sweta Raikundalia, Ling Ling Few, Siti Asma' Hassan, Get Bee Yvonne-Τee, Wei Cun See Too. Choline kinase and miR-32-5p: A crucial interaction promoting apoptosis and delaying wound repair in cervical cancer cells[J]. AIMS Biophysics, 2024, 11(3): 281-295. doi: 10.3934/biophy.2024016







 DownLoad:
DownLoad: