Changes in the structure and sizes of human and bovine serum albumins as well as polyvinyl alcohol macromolecules in aqueous solutions depending on temperature, concentration, and acid-base balance (pH) of the solutions are discussed. It is taken into consideration that the change in the hydrodynamic radius of a macromolecule is one of the indicators of structural phase transformations of globular proteins. The methods of the macromolecular radii determination from the shear viscosity and the self-diffusion of macromolecules in solutions are discussed. The hydrodynamic radius values of albumin and polyvinyl alcohol macromolecules obtained by the above methods are thoroughly compared. Consideration of these questions provides us with important information on the nature of the binding of water molecules with protein macromolecules.
Citation: Oleksii V. Khorolskyi, Nikolay P. Malomuzh. Macromolecular sizes of serum albumins in its aqueous solutions[J]. AIMS Biophysics, 2020, 7(4): 219-235. doi: 10.3934/biophy.2020017
Changes in the structure and sizes of human and bovine serum albumins as well as polyvinyl alcohol macromolecules in aqueous solutions depending on temperature, concentration, and acid-base balance (pH) of the solutions are discussed. It is taken into consideration that the change in the hydrodynamic radius of a macromolecule is one of the indicators of structural phase transformations of globular proteins. The methods of the macromolecular radii determination from the shear viscosity and the self-diffusion of macromolecules in solutions are discussed. The hydrodynamic radius values of albumin and polyvinyl alcohol macromolecules obtained by the above methods are thoroughly compared. Consideration of these questions provides us with important information on the nature of the binding of water molecules with protein macromolecules.

| [1] | Fenimore PW, Frauenfelder H, Magazù S, et al. (2013) Concepts and problems in protein dynamics. Chem Phys 424: 2-6. |
| [2] | Peters T (1996) All about Albumin: Biochemistry, Genetics, and Medical Applications New York: Academic Press, 1-432. |
| [3] | He XM, Carter DC (1992) Atomic structure and chemistry of human serum albumin. Nature 358: 209-215. |
| [4] | Majorek KA, Porebski PJ, Dayal A, et al. (2012) Structural and immunologic characterization of bovine, horse, and rabbit serum albumins. Mol Immunol 52: 174-182. |
| [5] | Akdogan Y, Reichenwallner J, Hinderberger D (2012) Evidence for water-tuned structural differences in proteins: an approach emphasizing variations in local hydrophilicity. PLoS ONE 7: e45681. |
| [6] | Carter DC, Ho JX (1994) Structure of serum albumin. Adv Protein Chem 45: 153-203. |
| [7] | Pagnotta SE, Ricci MA, Bruni F, et al. (2008) Water structure around trehalose. Chem Phys 345: 159-163. |
| [8] | Magazù S, Migliardo F, Benedetto A, et al. (2011) Thermal behavior of hydrated lysozyme in the presence of sucrose and trehalose by EINS. J Non-Cryst Solids 357: 664-670. |
| [9] | Luik AI, Naboka YuN, Mogilevich SE, et al. (1998) Study of human serum albumin structure by dynamic light scattering: two types of reactions under different pH and interaction with physiologically active compounds. Spectrochim Acta A Mol Biomol Spectrosc 54: 1503-1507. |
| [10] | Maslova MN, Zaritskiy AR, Chaykov LL (2014) The blood plasma particles sizes oscillations observed by dynamic light scattering. Biophys J 106: 457a-458a. |
| [11] | Magazù S, Maisano G, Migliardo P, et al. (1999) Experimental simulation of macromolecules in trehalose aqueous solutions: a photon correlation spectroscopy study. J Chem Phys 111: 9086-9092. |
| [12] | Sjöberg B, Mortensen K (1994) Interparticle interactions and structure in nonideal solutions of human serum albumin studied by small-angle neutron scattering and Monte Carlo simulation. Biophys Chemist 52: 131-138. |
| [13] | Kiselev MA, Gryzunov YA, Dobretsov GE, et al. (2001) The size of human serum albumin molecules in solution. Biophysics 46: 423-427. |
| [14] | Varga B, Migliardo F, Takacs E, et al. (2008) Neutron scattering studies on dUTPase complex in the presence of bioprotectant systems. Chem Phys 345: 250-258. |
| [15] | Chicea D, Chicea R, Chicea LM (2013) HSA particle size characterization by AFM. Rom Reports Phys 65: 178-185. |
| [16] | Olivieril JR, Craievich AF (1995) The subdomain structure of human serum albumin in solution under different pH conditions studied by small angle X-ray scattering. Eur Biophys J 24: 77-84. |
| [17] | Wilkins DK, Grimshaw SB, Receveur V, et al. (1999) Hydrodynamic radii of native and denatured proteins measured by pulse field gradient NMR techniques. Biochem 38: 16424-16431. |
| [18] | Magazù S (2000) NMR, static and dynamic light and neutron scattering investigations on polymeric aqueous solutions. J Mol Struct 523: 47-59. |
| [19] | Qian J, Tang Q, Cronin B, et al. (2008) Development of a high performance size exclusion chromatography method to determine the stability of human serum albumin in a lyophilized formulation of Interferon alfa-2b. J Chromatogr A 1194: 48-56. |
| [20] | Monkos K (2004) On the hydrodynamics and temperature dependence of the solution conformation of human serum albumin from viscometry approach. BBA-Proteins Proteom 1700: 27-34. |
| [21] | Einstein A (1906) Eine neue bestimmung der moleküldimensionen. Ann Phys 19: 289-305. |
| [22] | Batchelor GK (1967) An Introduction to Fluid Dynamics Cambridge: Cambridge university press, 1-615. |
| [23] | Malomuzh NP, Orlov EV (2005) Static shear viscosity of a bimodal suspension. Ukr J Phys 50: 618-622. |
| [24] | Orlov EV (2010) Shear viscosity of dispersions of particles with liquid shells. Colloid J 72: 820-824. |
| [25] | Khorolskyi OV (2018) Effective radii of macromolecules in dilute polyvinyl alcohol solutions. Ukr J Phys 63: 144-149. |
| [26] | Mooney RCL (1941) An X-ray study of the structure of polyvinyl alcohol. J Am Chem Soc 63: 2828-2832. |
| [27] | Bunn CW (1948) Crystal structure of polyvinyl alcohol. Nature 161: 929-930. |
| [28] | Colvin BG (1974) Crystal structure of polyvinyl alcohol. Nature 248: 756-759. |
| [29] | Gowariker VR, Viswanathan NV, Sreedhar J (1986) Polymer science New York: Wiley, 1-505. |
| [30] | Rumyantsev M, Gushchin A, Zelentsov S (2012) Effect of the type of hydrogen bonding on the reactivity of hydroxyl groups in the acetalization of poly(viny1 alcohol) with butanal. Polym Sci Series B 54: 464-471. |
| [31] | Karunakaran DJSA, Ganesh T, Sylvester MM, et al. (2014) Dielectric properties and analysis of H-bonded interaction study in complex systems of binary and ternary mixtures of polyvinyl alcohol with water and DMSO. Fluid Phase Equil 382: 300-306. |
| [32] | Tamai Y, Tanaka H, Nakanishi K (1996) Molecular dynamics study of polymer-water interaction in hydrogels. 1. Hydrogen-bond structure. Macromolecules 29: 6750-6760. |
| [33] | Budhlall BM, Landfester K, Sudol ED, et al. (2003) Characterization of partially hydrolyzed poly(vinyl alcohol). Effect of poly(vinyl alcohol) molecular architecture on aqueous phase conformation. Macromolecules 36: 9477-9484. |
| [34] | Fileti EE, Chaudhuri P, Canuto S (2004) Relative strength of hydrogen bond interaction in alcohol-water complexes. Chem Phys Lett 400: 494-499. |
| [35] | Pal S, Kundu TK (2012) Theoretical study of hydrogen bond formation in trimethylene glycol-water complex. Int Scholarly Res Notices 2012: 570394. |
| [36] | Rumyantsev M (2013) Influences of co-solvent on hydrogen bond reorganization in ternary poly(vinyl alcohol) solutions. Eur Polym J 49: 2257-2266. |
| [37] | Rumyantsev M, Zelentsov SV, Gushchin AV (2013) Retardation effect in acetalization of poly(vinyl alcohol) with butyraldehyde. Eur Polym J 49: 1698-1706. |
| [38] | Malomuzh NP, Bulavin LA, Gotsulskyi VYa, et al. (2020) Characteristic changes in the density and shear viscosity of human blood plasma with varying protein concentration. Ukr J Phys 65: 151-156. |
| [39] | Rumyantsev M, Rumyantsev S, Kazantsev OA, et al. (2020) Managing of hydrogen bonding in aqueous poly(vinyl alcohol) solutions: new perspectives towards preparation of more homogeneous poly(vinyl butyral). J Polym Res 27: 52. |
| [40] | Groda YG, Vikhrenko VS, Poghosyan AH, et al. (2015) Conformation and diffusion properties of polyvinyl alcohol and polyvinylpyrrolidone molecules. Elect J Nat Sci 24: 60-67. |
| [41] | Fisenko AI, Malomuzh NP (2009) To what extent is water responsible for the maintenance of the life for warm-blooded organisms? Int J Mol Sci 10: 2383-2411. |
| [42] | Bulavin LA, Malomuzh NP (2010) Dynamic phase transition in water as the most important factor of provision of denaturation of proteins in heat blood organisms. Phys Alive 18: 16-22. |
| [43] | Migliardo F, Magazù S, Caccamo MT (2013) Infrared, raman and INS studies of poly-ethylene oxide oligomers. J Mol Struct 1048: 261-266. |
| [44] | Caccamo MT, Magazù S (2016) Tagging the oligomer-to-polymer crossover on EG and PEGs by infrared and raman spectroscopies and by wavelet cross-correlation spectral analysis. Vibr Spectr 85: 222-227. |
| [45] | Garvey M, Tadros T, Vincent B (1974) A comparison of the volume occupied by macromolecules in the adsorbed state and in bulk solution: Adsorption of narrow molecular weight fractions of poly(vinyl alcohol) at the polystyrene/water interface. J Colloid Interf Sci 49: 57-68. |
| [46] | Wang B, Mukataka S, Kokufuta E, et al. (2000) Viscometric, light scattering, and size exclusion chromatography studies on the structural changes of aqueous poly(vinyl alcohol) induced by γ-ray irradiation. J Polym Sci Pol Phys 38: 214-221. |
| [47] | Wiśniewska M (2012) Temperature effects on the adsorption of polyvinyl alcohol on silica. Cent Eur J Chem 10: 1236-1244. |
| [48] | Khorolskyi OV, Moskalenko YuD (2020) Calculation of the macromolecular size of bovine serum albumin from the viscosity of its aqueous solutions. Ukr J Phys 65: 41-49. |
| [49] | Magazù S, Calabrò E (2011) Studying the electromagnetic-induced changes of the secondary structure of bovine serum albumin and the bioprotective effectiveness of trehalose by Fourier transform infrared spectroscopy. J Phys Chem B 115: 6818-6826. |
| [50] | Magazù S, Calabrò E, Campo S, et al. (2012) New insights into bioprotective effectiveness of disaccharides: an FTIR study of human haemoglobin aqueous solutions exposed to static magnetic fields. J Bio Phys 38: 61-74. |
| [51] | Minutoli L, Altavilla D, Bitto A, et al. (2008) Trehalose: a biophysics approach to modulate the inflammatory response during endotoxic shock. Eur J Pharm 589: 272-280. |
| [52] | Yadav S, Shire SJ, Kalonia DS (2011) Viscosity analysis of high concentration bovine serum albumin aqueous solutions. Pharm Res 28: 1973-1983. |
| [53] | Gaigalas AK, Hubbard JB, McCurley M, et al. (1992) Diffusion of bovine serum albumin in aqueous solutions. J Phys Chem 96: 2355-2359. |
| [54] | Keller KH, Canales ER, Yum SI (1971) Tracer and mutual diffusion coefficients of proteins. J Phys Chem 75: 379-387. |
| [55] | Jachimska B, Pajor A (2017) Physico-chemical characterization of bovine serum albumin in solution and as deposited on surfaces. Bioelectrochem 87: 138-146. |
| [56] | Brenner H (1974) Rheology of a dilute suspension of axisymmetric brownian particles. J Multiphase Flow 1: 195-341. |
| [57] | Khorolskyi OV (2019) Calculation of the effective macromolecular radii of human serum albumin from the shear viscosity data for its aqueous solutions. Ukr J Phys 64: 287-292. |
| [58] | Bardik V, Gotsulskii V, Pavlov E, et al. (2012) Light scattering study of human serum albumin in pre-denaturation: relation to dynamic transition in water at 42 °C. J Mol Liq 176: 60-64. |
| [59] | Atamas N, Bardik V, Bannikova A, et al. (2017) The effect of water dynamics on conformation changes of albumin in pre-denaturation state: photon correlation spectroscopy and simulation. J Mol Liq 235: 17-23. |
| [60] | Atamas N, Bardik VY, Komisarenko S, et al. (2019) Water dynamics and stability of major blood proteins at pre-denaturation stage. Atti Accad Pelorit Pericol Cl Sci Fis Mat Nat 97: A16. |
| [61] | Hushcha TO, Luik AI, Naboka YN (2000) Conformation changes of albumin in its interaction with physiologically active compounds as studied by quasi-elastic light scattering spectroscopy and ultrasonic method. Talanta 53: 29-34. |
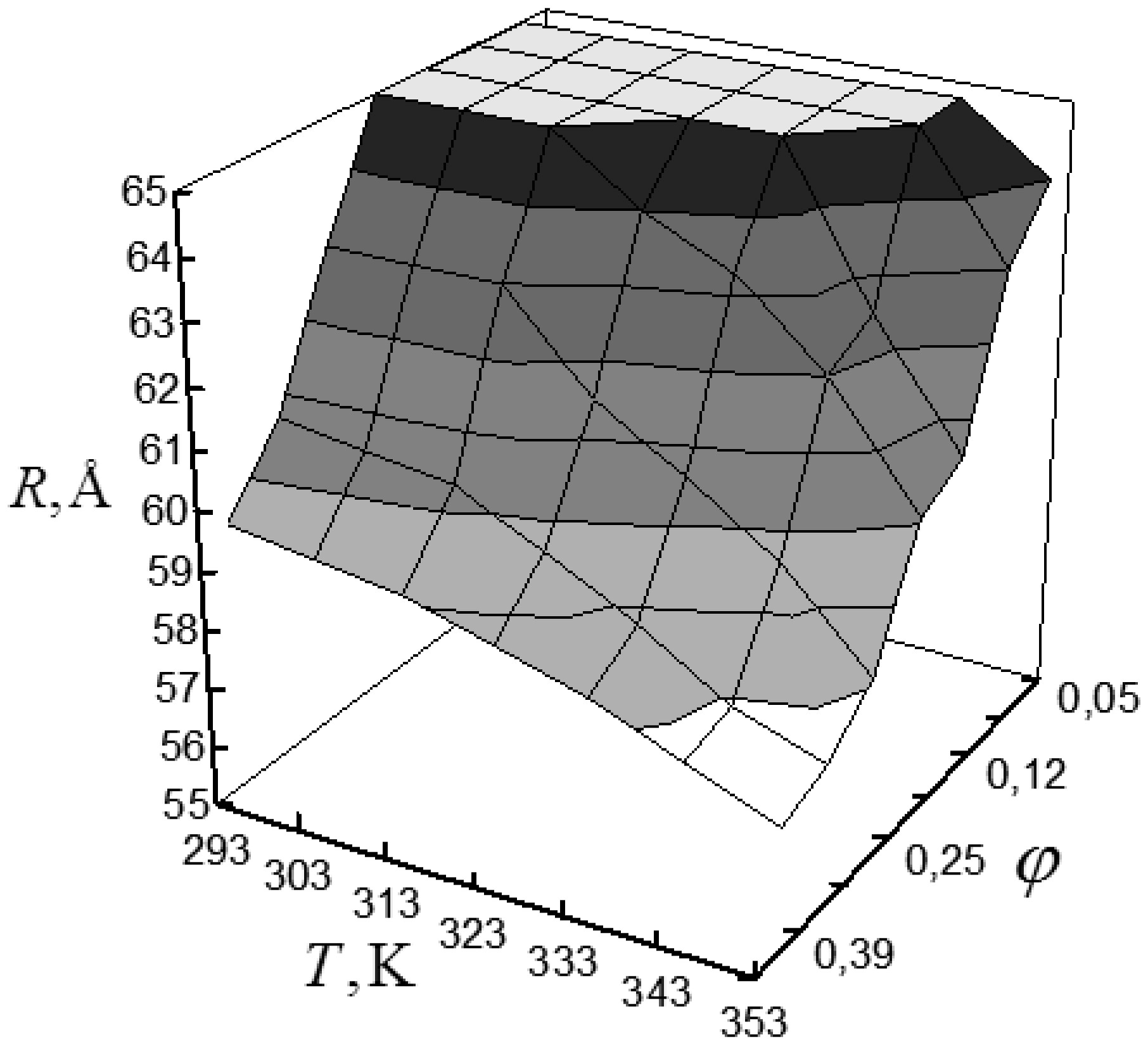
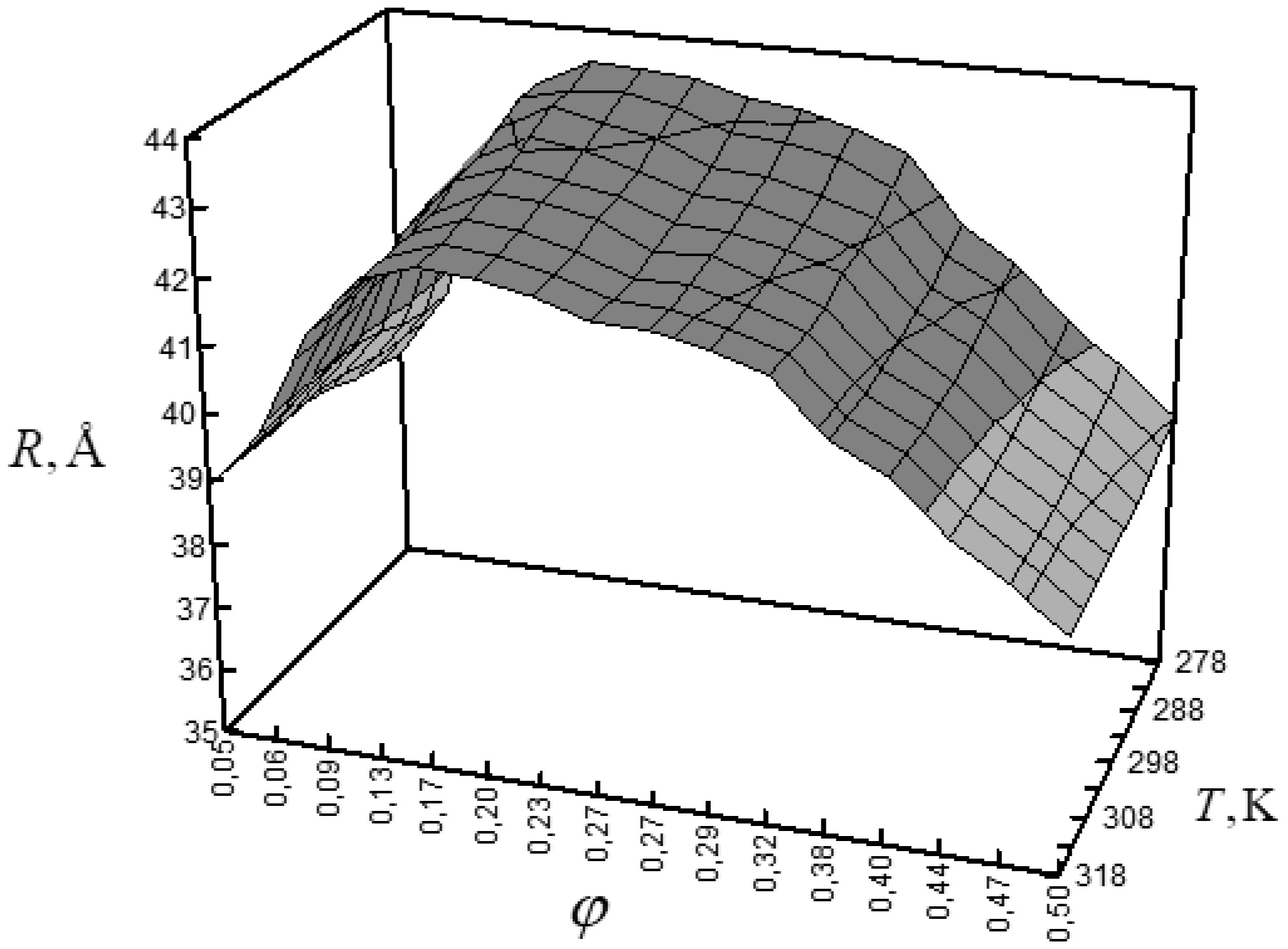
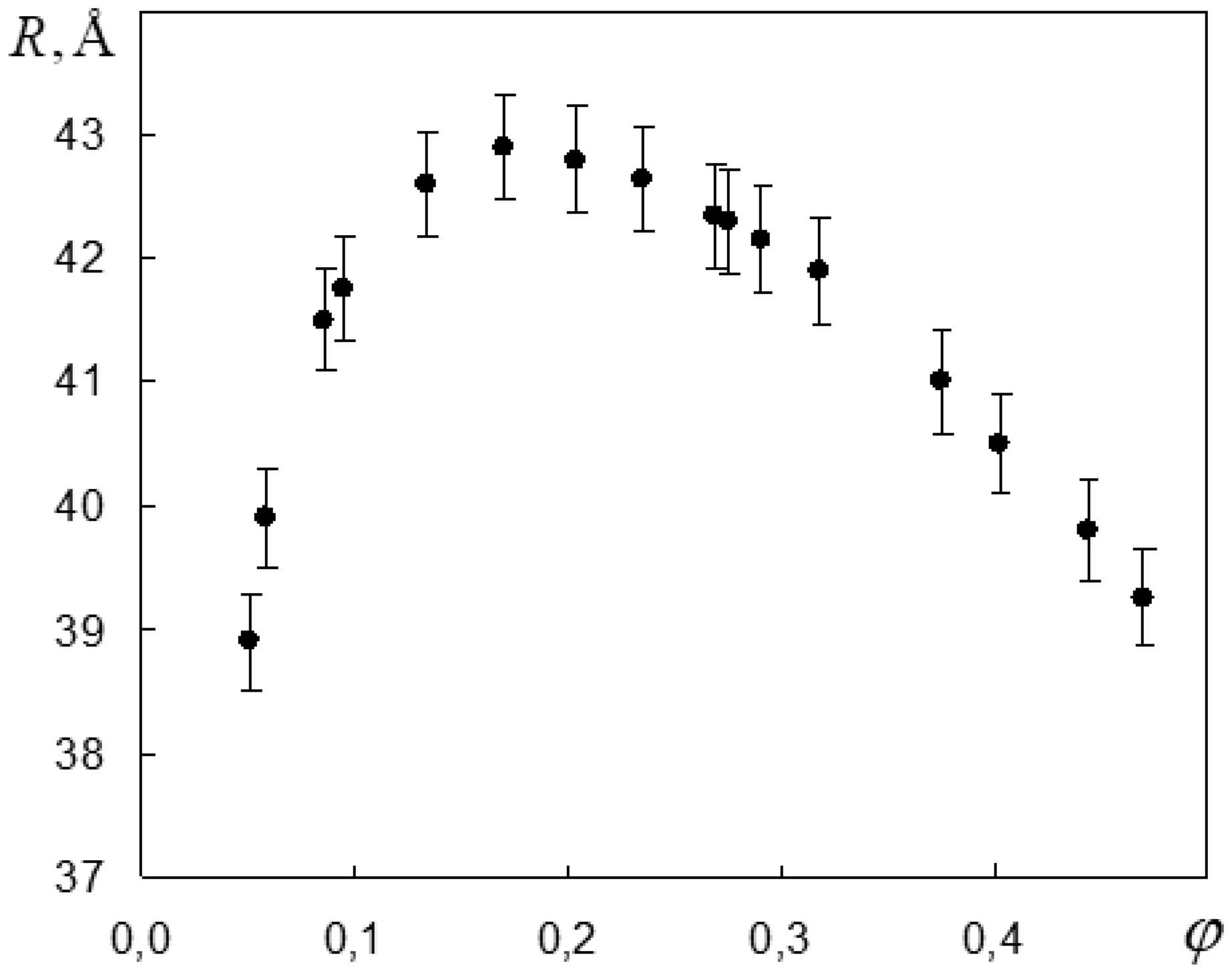
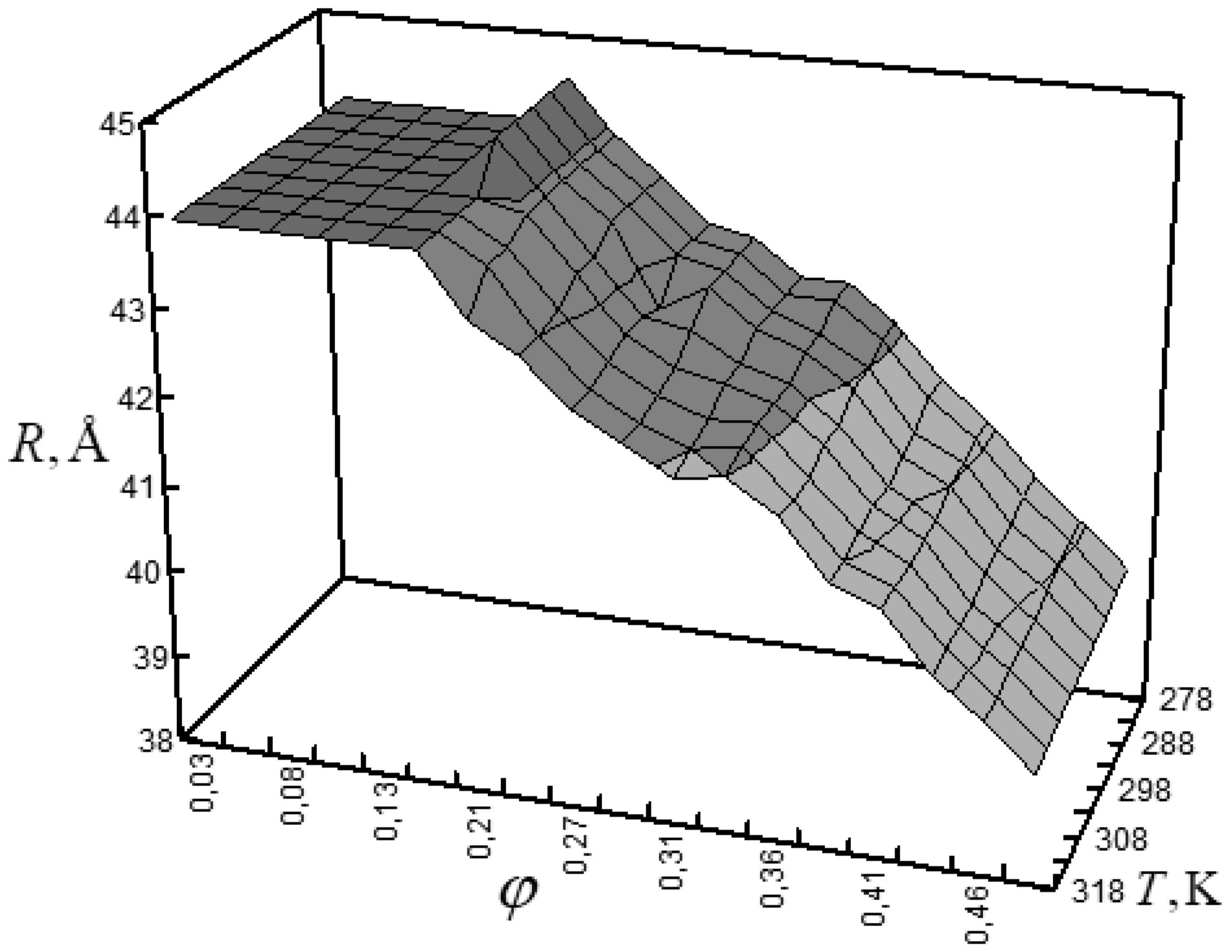
Figures(6) / Tables(1)

Oleksii V. Khorolskyi, Nikolay P. Malomuzh. Macromolecular sizes of serum albumins in its aqueous solutions[J]. AIMS Biophysics, 2020, 7(4): 219-235. doi: 10.3934/biophy.2020017







 DownLoad:
DownLoad: