Marine algae are a valuable source of sulfated polysaccharides presenting varied structural characteristics and biological activities. Regarding sulfated polysaccharides extracted from marine microalgae, molecular characteristics and bioactivity have yet to be fully explored, especially in diatoms. Sulfated polysaccharides from marine diatoms have great potential to produce numerous health benefits and lead to new biomedical materials. Nevertheless, these potential applications are based on the polysaccharide molecular characteristics, which define their functional properties. Therefore, a detailed understanding of sulfated polysaccharides from marine diatoms may represent the starting point for a broad development of innovative applications, especially in the biomedical area. In this context, the present manuscript aims to review marine diatom sulfated polysaccharides' molecular characteristics and biological activity, looking for a more profound knowledge of these macromolecules and their potential applications.
Citation: Valeria Miranda-Arizmendi, Diana Fimbres-Olivarria, Anselmo Miranda-Baeza, Agustín Rascón-Chu, Jorge Marquez-Escalante, Jaime Lizardi-Mendoza, Mayra A. Méndez-Encinas, Elizabeth Carvajal-Millan. Sulfated polysaccharides from marine diatoms: Insight into molecular characteristics and biological activity[J]. AIMS Bioengineering, 2024, 11(1): 110-129. doi: 10.3934/bioeng.2024007
Marine algae are a valuable source of sulfated polysaccharides presenting varied structural characteristics and biological activities. Regarding sulfated polysaccharides extracted from marine microalgae, molecular characteristics and bioactivity have yet to be fully explored, especially in diatoms. Sulfated polysaccharides from marine diatoms have great potential to produce numerous health benefits and lead to new biomedical materials. Nevertheless, these potential applications are based on the polysaccharide molecular characteristics, which define their functional properties. Therefore, a detailed understanding of sulfated polysaccharides from marine diatoms may represent the starting point for a broad development of innovative applications, especially in the biomedical area. In this context, the present manuscript aims to review marine diatom sulfated polysaccharides' molecular characteristics and biological activity, looking for a more profound knowledge of these macromolecules and their potential applications.

| [1] |
Lordan S, Ross RP, Stanton C (2011) Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Marine Drugs 9: 1056-1100. https://doi.org/10.3390/md9061056 
|
| [2] |
Ntie-Kang F, Svozil D (2020) An enumeration of natural products from microbial, marine and terrestrial sources. Physical Sciences Reviews 5: 20180121. https://doi.org/10.1515/psr-2018-0121
|
| [3] |
Hamed I, Özogul F, Özogul Y, et al. (2015) Marine bioactive compounds and their health benefits: A review. Compr Rev Food Sci F 14: 446-465. https://doi.org/10.1111/1541-4337.12136
|
| [4] |
Pangestuti R, Kim S-K (2015) An overview of phycocolloids: The principal commercial seaweed extracts. Marine Algae Extracts: Processes, Products, and Applications . Germany: Wiley-VCH 319-330. https://doi.org/10.1002/9783527679577.ch19
|
| [5] |
Khan T, Date A, Chawda H, et al. (2019) Polysaccharides as potential anticancer agents—A review of their progress. Carbohyd Polym 210: 412-428. https://doi.org/10.1016/j.carbpol.2019.01.064
|
| [6] |
Sun T, Tao H, Xie J, et al. (2010) Degradation and antioxidant activity of κ-carrageenans. J Appl Polym Sci 117: 194-199. https://doi.org/10.1002/app.31955
|
| [7] |
Wijesekara I, Pangestuti R, Kim S-K (2011) Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohyd Polym 84: 14-21. https://doi.org/10.1016/j.carbpol.2010.10.062
|
| [8] |
de Jesus Raposo M, de Morais A, de Morais R (2015) Marine polysaccharides from algae with potential biomedical applications. Marine Drugs 13: 2967-3028. https://doi.org/10.3390/md13052967
|
| [9] |
Prybylski N, Toucheteau C, El Alaoui H, et al. (2020) Bioactive polysaccharides from microalgae. Handbook of Microalgae-Based Processes and Products . Amsterdam: Elsevier 533-571. https://doi.org/10.1016/B978-0-12-818536-0.00020-8
|
| [10] | McKeen LW (2013) The Effect of UV Light and Weather on Plastics and Elastomers. Amsterdam: Elsevier. https://doi.org/10.1016/C2011-0-07329-5 |
| [11] | Rio AR del, Ramírez-Gilly M, Tecante A (2017) Flow properties of lambda carrageenan in aqueous systems. Biological Activities and Application of Marine Polysaccharides . London: InTech. https://doi.org/10.5772/65785 |
| [12] |
Danaei M, Dehghankhold M, Ataei S, et al. (2018) Impact of particle size and polydispersity index on the clinical applications of lipidic nanocarrier systems. Pharmaceutics 10: 57. https://doi.org/10.3390/pharmaceutics10020057
|
| [13] | Guo MQ, Hu X, Wang C, et al. (2017) Polysaccharides: Structure and solubility. Solubility of Polysaccharides . London: InTech 8-21. https://doi.org/10.5772/intechopen.71570 |
| [14] |
Burchard W (1999) Solution properties of branched macromolecules. Branched Polymers II . Berlin: Springer 113-194. https://doi.org/10.1007/3-540-49780-3_3
|
| [15] |
Tande BM, Wagner NJ, Mackay ME, et al. (2001) Viscosimetric, hydrodynamic, and conformational properties of dendrimers and dendrons. Macromolecules 34: 8580-8585. https://doi.org/10.1021/ma011265g
|
| [16] |
Teraoka I (2002) Polymer Solutions: An Introduction to Physical Properties. New York: Wiley. https://doi.org/10.1002/0471224510
|
| [17] |
Cardozo KHM, Guaratini T, Barros MP, et al. (2007) Metabolites from algae with economical impact. Comp Biochem Physiol C: Toxicol Pharm 146: 60-78. https://doi.org/10.1016/j.cbpc.2006.05.007
|
| [18] |
Pangestuti R, Kim S-K (2014) Biological Activities of Carrageenan. Advances in Food and Nutrition Research . Amsterdam: Elsevier 113-124. https://doi.org/10.1016/B978-0-12-800269-8.00007-5
|
| [19] | Venugopal V (2008) Marine products for healthcare: functional and bioactive nutraceutical compounds from the ocean. Marine Products for Healthcare . Boca Raton: CRC Press. https://doi.org/10.1201/9781420052640 |
| [20] |
Rengasamy KRR, Mahomoodally MF, Aumeeruddy MZ, et al. (2020) Bioactive compounds in seaweeds: An overview of their biological properties and safety. Food Chem Toxicol 135: 111013. https://doi.org/10.1016/j.fct.2019.111013
|
| [21] |
Tseng CK (2001) Algal biotechnology industries and research activities in china. J Appl Phycol 13: 375-380. http://dx.doi.org/10.1023/A:1017972812576
|
| [22] | Kraan S (2012) Algal polysaccharides, novel applications and outlook. Carbohydrates - Comprehensive Studies on Glycobiology and Glycotechnology . London: InTech. https://doi.org/10.1016/B978-0-12-812360-7.00005-7 |
| [23] |
Zaporozhets TS, Besednova NN, Kuznetsova TA, et al. (2014) The prebiotic potential of polysaccharides and extracts of seaweeds. Russ J Mar Biol 40: 1-9. https://doi.org/10.1134/S1063074014010106
|
| [24] |
Kulkarni VS, Shaw C (2016) Use of polymers and thickeners in semisolid and liquid formulations. Essential Chemistry for Formulators of Semisolid and Liquid Dosages . New York: Academic Press 43-69. https://doi.org/10.1016/B978-0-12-801024-2.00005-4
|
| [25] |
Ale MT, Mikkelsen JD, Meyer AS (2011) Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar Drugs 9: 2106-2130. https://doi.org/10.3390/md9102106
|
| [26] |
Kidgell JT, Magnusson M, de Nys R, et al. (2019) Ulvan: A systematic review of extraction, composition and function. Algal Res 39: 101422. https://doi.org/10.1016/j.algal.2019.101422
|
| [27] |
Raposo M, De Morais R, Bernardo de Morais A (2013) Bioactivity and applications of sulphated polysaccharides from marine microalgae. Mar Drugs 11: 233-252. https://doi.org/10.3390/md11010233
|
| [28] |
Singh P, Maparu AK, Shah S, et al. (2021) Biomimetic algal polysaccharide coated 3D nanofibrous scaffolds promote skin extracellular matrix formation. Mat Sci Eng C 119: 111580. https://doi.org/10.1016/j.msec.2020.111580
|
| [29] |
Vuppaladadiyam AK, Prinsen P, Raheem A, et al. (2018) Microalgae cultivation and metabolites production: a comprehensive review. Biofuel Bioprod Bior 12: 304-324. https://doi.org/10.1002/bbb.1864
|
| [30] |
Gügi B, Le Costaouec T, Burel C, et al. (2015) Diatom-specific oligosaccharide and polysaccharide structures help to unravel biosynthetic capabilities in diatoms. Mar Drugs 13: 5993-6018. https://doi.org/10.3390/md13095993
|
| [31] | Hoek C, Mann DG, Jahns HM (1995) Algae: an Introduction to Phycology. New York: Cambridge University Press. |
| [32] |
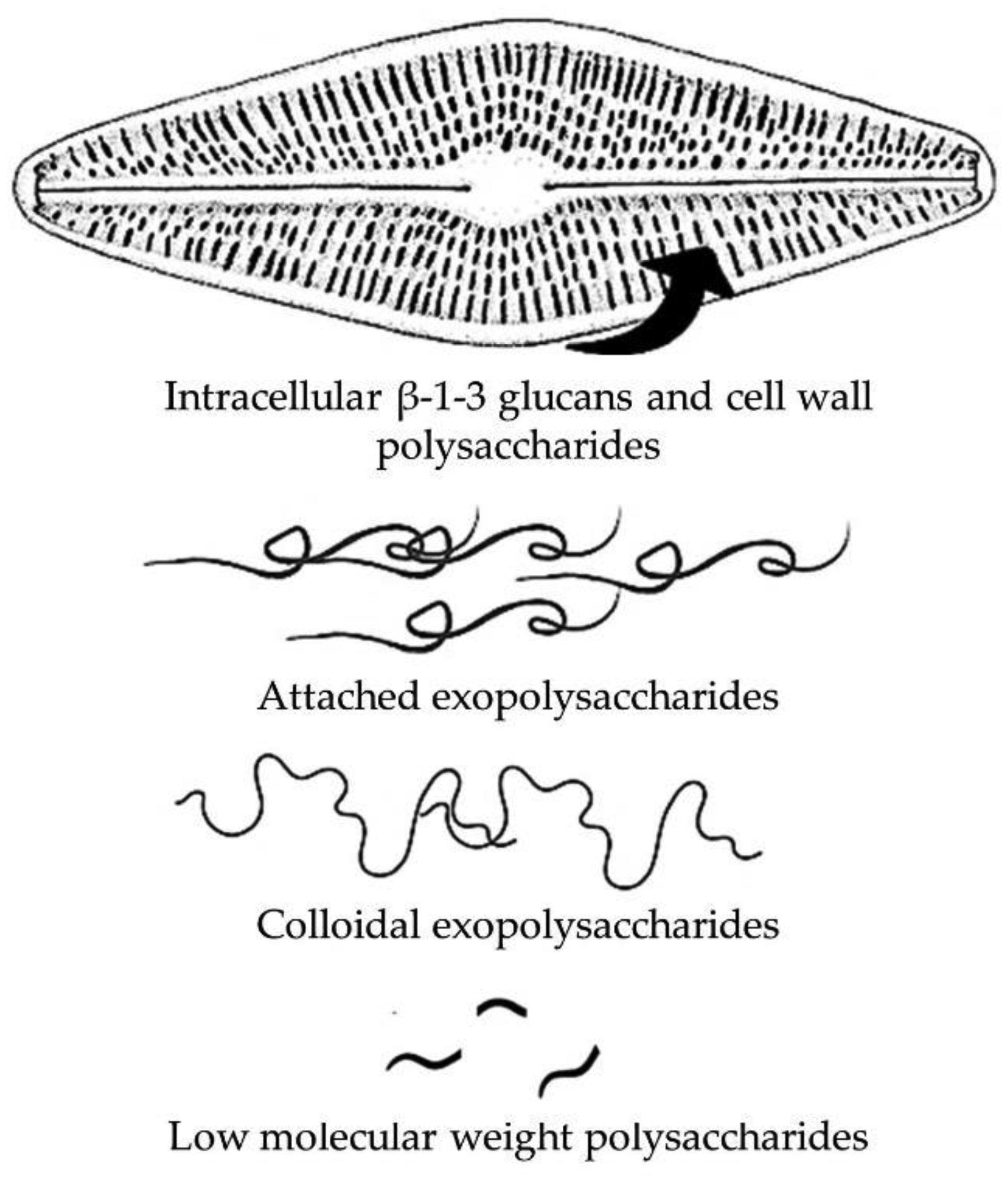
Shniukova EI, Zolotareva EK (2015) Diatom exopolysaccharides: a review. Inter J Algae 17: 50-67. https://doi.org/10.1615/InterJAlgae.v17.i1.50
|
| [33] |
Alipour L, Hamamoto M, Nakashima S, et al. (2016) Infrared microspectroscopy of bionanomaterials (diatoms) with careful evaluation of void effects. Appl Spectrosc 70: 427-442. https://doi.org/10.1177/0003702815626665
|
| [34] |
Underwood GJC, Paterson DM (2003) The importance of extracellular carbohydrate productionby marine epipelic diatoms. Adv Bot Res 40: 183-240. https://doi.org/10.1016/S0065-2296(05)40005-1
|
| [35] |
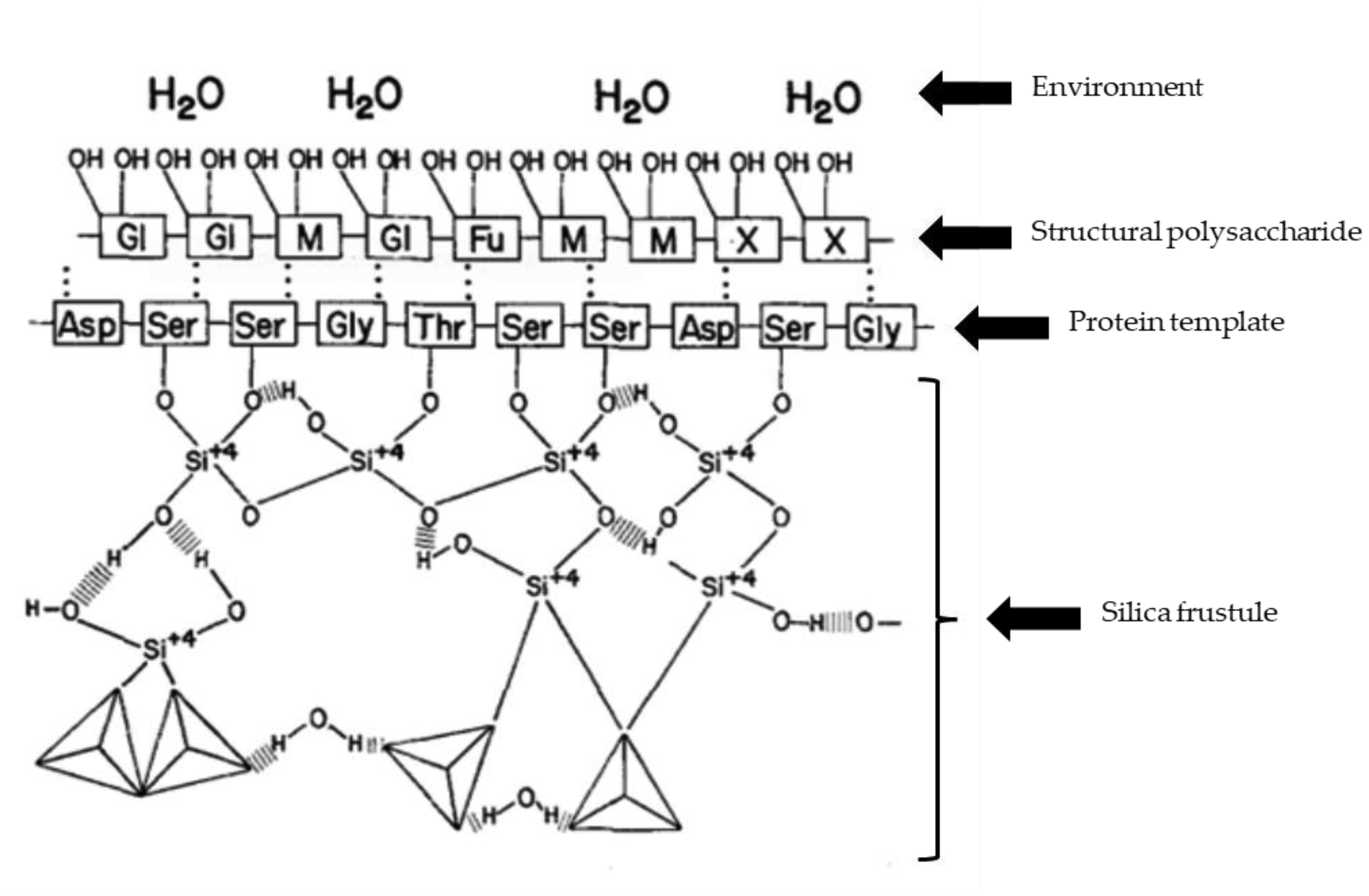
Hecky RE, Mopper K, Kilham P, et al. (1973) The amino acid and sugar composition of diatom cell-walls. Mar Biol 19: 323-331. https://doi.org/10.1007/BF00348902
|
| [36] | Decho AW (1990) Microbial exopolymer secretions in ocean environments: their role (s) in food webs and marine processes. Oceanography and Marine Biology: An Annual Review . London: Aberdeen University Press 73-153. |
| [37] |
Orvain F, Galois R, Barnard C, et al. (2003) Carbohydrate production in relation to microphytobenthic biofilm development: An integrated approach in a tidal mesocosm. Microb Ecol 45: 237-251. https://doi.org/10.1007/s00248-002-2027-7
|
| [38] |
Dai WY, Han L, Li PF, et al. (2023) The sulfate assimilation and reduction of marine microalgae and the regulation of illumination. Mar Environ Res 191: 106156. https://doi.org/10.1016/j.marenvres.2023.106156
|
| [39] |
Takahashi H, Kopriva S, Giordano M, et al. (2011) Sulfur assimilation in photosynthetic organisms: Molecular functions and regulations of transporters and assimilatory enzymes. Annu Rev Plant Biol 62: 157-184. https://doi.org/10.1146/annurev-arplant-042110-103921
|
| [40] |
Takahashi H, Buchner P, Yoshimoto N, et al. (2012) Evolutionary relationships and functional diversity of plant sulfate transporters. Front Plant Sci 2: 119. https://doi.org/10.3389/fpls.2011.00119
|
| [41] |
Buendía-Flores M, Tavera R, Novelo E (2015) Florística y ecología de diatomeas bentónicas de la zona lacustre de Xochimilco-Tláhuac, México. Bot Sci 93: 531-558. https://doi.org/10.17129/botsci.148
|
| [42] |
Lee RE (2018) Phycology. New York: Cambridge University Press.
|
| [43] |
Miranda-Arizmendi V, Fimbres-Olivarria D, Miranda-Baeza A, et al. (2022) Sulfated polysaccharides from chaetoceros muelleri: Macromolecular characteristics and bioactive properties. Biology 11: 1476. https://doi.org/10.3390/biology11101476
|
| [44] |
Geresh S, Mamontov A, Weinstein J (2002) Sulfation of extracellular polysaccharides of red microalgae: Preparation, characterization and properties. J Biochem Bioph Meth 50: 179-187. https://doi.org/10.1016/S0165-022X(01)00185-3
|
| [45] |
Jiang JL, Zhang WZ, Ni WX, et al. (2021) Insight on structure-property relationships of carrageenan from marine red algal: A review. Carbohyd Polym 257: 117642. https://doi.org/10.1016/j.carbpol.2021.117642
|
| [46] | Wang J, Hu S, Nie S, et al. (2016) Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxid Med Cell Longev 2016: 1-13. https://doi.org/10.1155/2016/5692852 |
| [47] |
Kang MC, Kim SY, Kim EA, et al. (2015) Antioxidant activity of polysaccharide purified from acanthopanax koreanum nakai stems in vitro and in vivo zebrafish model. Carbohyd Polym 127: 38-46. https://doi.org/10.1016/j.carbpol.2015.02.043
|
| [48] |
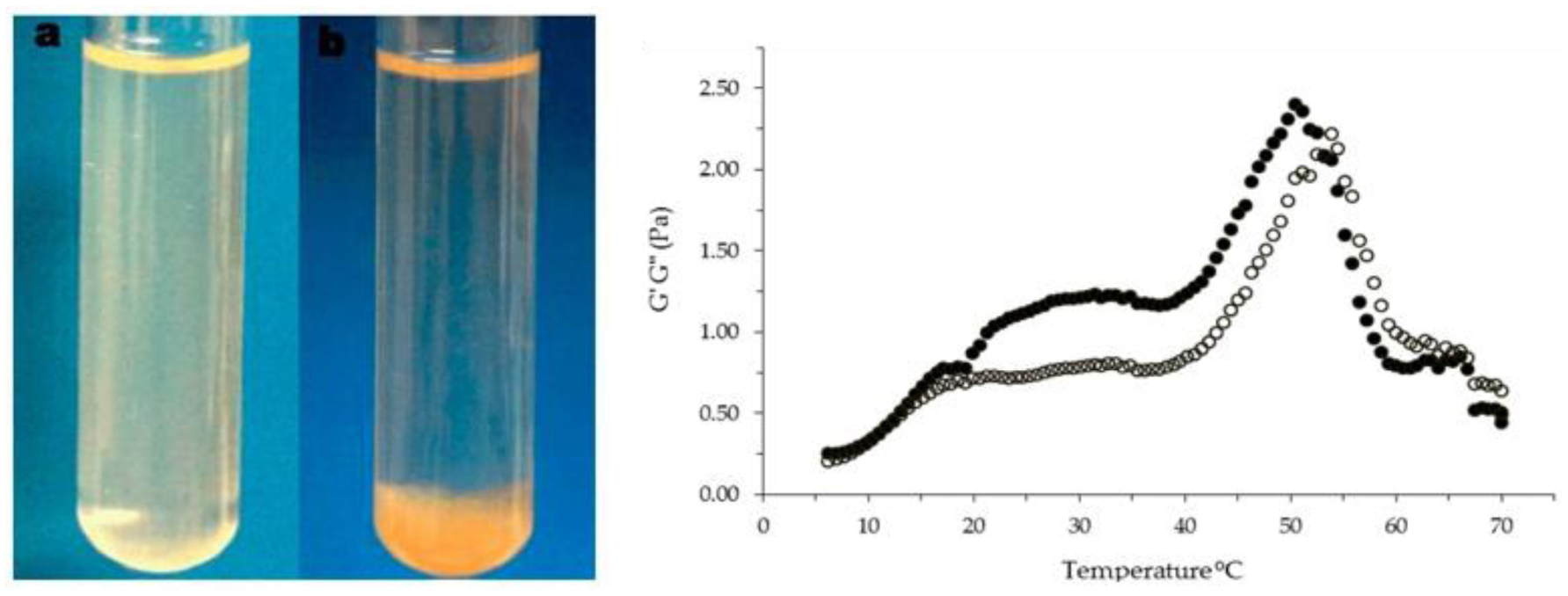
Fimbres-Olivarría D, López-Elías J, Carvajal-Millán E, et al. (2016) Navicula sp. sulfated polysaccharide gels induced by Fe(III): Rheology and microstructure. Int J Mol Sci 17: 1238. https://doi.org/10.3390/ijms17081238
|
| [49] |
González-Vega RI, Del-Toro-Sánchez CL, Moreno-Corral RA, et al. (2022) Sulfated polysaccharide-rich extract from Navicula incerta: Physicochemical characteristics, antioxidant activity, and anti-hemolytic property. AIMS Bioeng 9: 364-382. https://doi.org/10.3934/bioeng.2022027
|
| [50] |
Xia S, Gao B, Li A, et al. (2014) Preliminary characterization, antioxidant properties and production of chrysolaminarin from marine diatom odontella aurita. Mar Drugs 12: 4883-4897. https://doi.org/10.3390/md12094883
|
| [51] |
Lim S, Choi J, Park H (2015) Antioxidant activities of fucoidan degraded by gamma irradiation and acidic hydrolysis. Radiat Phys Chem 109: 23-26. https://doi.org/10.1016/j.radphyschem.2014.12.008
|
| [52] |
Gómez-Ordóñez E, Jiménez-Escrig A, Rupérez P (2014) Bioactivity of sulfated polysaccharides from the edible red seaweed Mastocarpus stellatus. Bioact Carbohyd Dietary Fibre 3: 29-40. https://doi.org/10.1016/j.bcdf.2014.01.002
|
| [53] |
Wang J, Zhang Q, Zhang Z, et al. (2008) Antioxidant activity of sulfated polysaccharide fractions extracted from laminaria japonica. Int J Biol Macromol 42: 127-132. https://doi.org/10.1016/j.ijbiomac.2007.10.003
|
| [54] |
Kusaikin MI, Ermakova SP, Shevchenko NM, et al. (2010) Structural characteristics and antitumor activity of a new chrysolaminaran from the diatom alga Synedra acus. Chem Nat Compd 46: 1-4. https://doi.org/10.1007/s10600-010-9510-z
|
| [55] |
Yang S, Wan H, Wang R, et al. (2019) Sulfated polysaccharides from Phaeodactylum tricornutum: isolation, structural characteristics, and inhibiting HepG2 growth activity in vitro. PeerJ 7: e6409. https://doi.org/10.7717/peerj.6409
|
| [56] |
Lee JB, Hayashi K, Hirata M, et al. (2006) Antiviral sulfated polysaccharide from Navicula directa, a diatom collected from deep-sea water in Toyama Bay. Biol Pharm Bull 29: 2135-2139. https://doi.org/10.1248/bpb.29.2135
|
| [57] |
Yim JH, Kim SJ, Ahn SH, et al. (2004) Antiviral effects of sulfated exopolysaccharide from the marine microalga gyrodinium impudicum strain KG03. Mar Biotechnol 6: 17-25. https://doi.org/10.1007/s10126-003-0002-z
|
| [58] |
Rizzi J, Moro TR, Winnischofer SMB, et al. (2023) Chemical structure and biological activity of the (1 → 3)-linked β-D-glucan isolated from marine diatom Conticribra weissflogii. Int J Biol Macromol 224: 584-593. https://doi.org/10.1016/j.ijbiomac.2022.10.147
|
| [59] |
Lai HL, Yang LC, Lin PT, et al. (2020) Phagocytosis activity of three sulfated polysaccharides purified from a marine diatom cultured in a semi-continuous system. Int J Biol Macromols 155: 951-960. https://doi.org/10.1016/j.ijbiomac.2019.11.054
|
| [60] |
Beaumont M, Tran R, Vera G, et al. (2021) Hydrogel-forming algae polysaccharides: From seaweed to biomedical applications. Biomacromolecules 22: 1027-1052. https://doi.org/10.1021/acs.biomac.0c01406
|
| [61] |
Guo R, Chen M, Ding Y, et al. (2022) Polysaccharides as potential anti-tumor biomacromolecules—a review. Front Nutr 9: 838179. https://doi.org/10.3389/fnut.2022.838179
|
| [62] |
Copeland R, Balasubramaniam A, Tiwari V, et al. (2008) Using a 3-O-sulfated heparin octasaccharide to inhibit the entry of herpes simplex virus type 1. Biochemistry 47: 5774-5783. https://doi.org/10.1021/bi800205t
|
| [63] |
Kwon PS, Oh H, Kwon S-J, et al. (2020) Sulfated polysaccharides effectively inhibit SARS-CoV-2 in vitro. Cell Discov 6: 50. https://doi.org/10.1038/s41421-020-00192-8
|
| [64] |
Morokutti-Kurz M, Fröba M, Graf P, et al. (2021) Iota-carrageenan neutralizes SARS-CoV-2 and inhibits viral replication in vitro. PLoS One 16: e0237480. https://doi.org/10.1371/journal.pone.0237480
|
| [65] |
Song S, Peng H, Wang Q, et al. (2020) Inhibitory activities of marine sulfated polysaccharides against SARS-CoV-2. Food Funct 11: 7415-7420. https://doi.org/10.1039/D0FO02017F
|
| [66] |
Marks RM, Lu H, Sundaresan R, et al. (2001) Probing the interaction of dengue virus envelope protein with heparin: Assessment of glycosaminoglycan-derived inhibitors. J Med Chem 44: 2178-2187. https://doi.org/10.1021/jm000412i
|
| [67] |
Panggabean JA, Adiguna SP, Rahmawati SI, et al. (2022) Antiviral activities of algal-based sulfated polysaccharides. Molecules 27: 1178. https://doi.org/10.3390/molecules27041178
|
| [68] |
Huang L, Shen M, Morris GA, et al. (2019) Sulfated polysaccharides: Immunomodulation and signaling mechanisms. Trends Food Sci Tech 92: 1-11. https://doi.org/10.1016/j.tifs.2019.08.008
|
| [69] |
Schepetkin IA, Faulkner CL, Nelson-Overton LK, et al. (2005) Macrophage immunomodulatory activity of polysaccharides isolated from Juniperus scopolorum. Int Immunopharmacol 5: 1783-1799. https://doi.org/10.1016/j.intimp.2005.05.009
|
| [70] |
Khil'chenko SR, Zaporozhets TS, Shevchenko NM, et al. (2011) Immunostimulatory activity of fucoidan from the brown alga fucus evanescens: Role of sulfates and acetates. J Carbohyd Chem 30: 291-305. https://doi.org/10.1080/07328303.2011.604456
|
| [71] |
Ohta Y, Lee JB, Hayashi K, et al. (2007) In vivo anti-influenza virus activity of an immunomodulatory acidic polysaccharide isolated from cordyceps militaris grown on germinated soybeans. J Agric Food Chem 55: 10194-10199. https://doi.org/10.1021/jf0721287
|
| [72] |
Xie G, Schepetkin IA, Siemsen DW, et al. (2008) Fractionation and characterization of biologically-active polysaccharides from Artemisia tripartita. Phytochemistry 69: 1359-1371. https://doi.org/10.1016/j.phytochem.2008.01.009
|
| [73] |
Xu X, Yan H, Zhang X (2012) Structure and immuno-stimulating activities of a new heteropolysaccharide from lentinula edodes. J Agric Food Chem 60: 11560-11566. https://doi.org/10.1021/jf304364c
|
| [74] |
Ramesh H (2002) Effect of fenugreek (Trigonella foenum-graecum L.) galactomannan fractions on phagocytosis in rat macrophages and on proliferation and IgM secretion in HB4C5 cells. Carbohyd Polym 50: 79-83. https://doi.org/10.1016/S0144-8617(01)00377-0
|
| [75] |
Engel A, Thoms S, Riebesell U, et al. (2004) Polysaccharide aggregation as a potential sink of marine dissolved organic carbon. Nature 428: 929-932. https://doi.org/10.1038/nature02453
|
| [76] |
Chin WC, Orellana MV, Verdugo P (1998) Spontaneous assembly of marine dissolved organic matter into polymer gels. Nature 391: 568-572. https://doi.org/10.1038/35345
|
| [77] |
Vidal-Melgosa S, Sichert A, Francis TB, et al. (2021) Diatom fucan polysaccharide precipitates carbon during algal blooms. Nat Commun 12: 1150. https://doi.org/10.1038/s41467-021-21009-6
|
| [78] |
Bernaerts TMM, Kyomugasho C, Van Looveren N, et al. (2018) Molecular and rheological characterization of different cell wall fractions of Porphyridium cruentum. Carbohyd Polym 195: 542-550. https://doi.org/10.1016/j.carbpol.2018.05.001
|
| [79] |
Costa JAV, Lucas BF, Alvarenga AGP, et al. (2021) Microalgae polysaccharides: An overview of production, characterization, and potential applications. Polysaccharides 2: 759-772. https://doi.org/10.3390/polysaccharides2040046
|
| [80] |
Malkin AY, Isayev AI (2006) Concepts, methods, and applications. Appl Rheol 16: 240-241. https://doi.org/10.1515/arh-2006-0039
|
| [81] |
Fernandes HL, Lupi F, Tomé MM, et al. (1991) Rheological behaviour of the culture medium during growth of the microalga Botryococcus braunii. Bioresource Technol 38: 133-136. https://doi.org/10.1016/0960-8524(91)90144-9
|
| [82] |
Cho M, Yang C, Kim SM, et al. (2010) Molecular characterization and biological activities of watersoluble sulfated polysaccharides from Enteromorpha prolifera. Food Sci Biotechnol 19: 525-533. https://doi.org/10.1007/s10068-010-0073-3
|
| [83] |
Liang Y, He J, Guo B (2021) Functional hydrogels as wound dressing to enhance wound healing. ACS Nano 15: 12687-12722. https://doi.org/10.1021/acsnano.1c04206
|
| [84] |
Running CA, Falshaw R, Janaswamy S (2012) Trivalent iron induced gelation in lambda-carrageenan. Carbohydrate Polym 87: 2735-2739. https://doi.org/10.1016/j.carbpol.2011.11.018
|
| [85] |
Tiwari A, Melchor-Martínez EM, Saxena A, et al. (2021) Therapeutic attributes and applied aspects of biological macromolecules (polypeptides, fucoxanthin, sterols, fatty acids, polysaccharides, and polyphenols) from diatoms — A review. Int J Biol Macromol 171: 398-413. https://doi.org/10.1016/j.ijbiomac.2020.12.219
|
| [86] |
Moreira JB, Vaz BDS, Cardias BB, et al. (2022) Microalgae polysaccharides: An alternative source for food production and sustainable agriculture. Polysaccharides 3: 441-457. https://doi.org/10.3390/polysaccharides3020027
|
| [87] |
Apostolova E, Lukova P, Baldzhieva A, et al. (2020) Immunomodulatory and anti-inflammatory effects of fucoidan: A review. Polymers 12: 2338. https://doi.org/10.3390/polym12102338
|
| [88] |
Zayed A, Cao HTT, Trang VTD, et al. (2023) Structural tailoring of fucoidan backbones for maximizing their benefits: Enzymatic, chemical, and physical strategies. J Appl Phycol 35: 2445-2462. https://doi.org/10.1007/s10811-023-03036-6
|
Figures(3) / Tables(2)

Valeria Miranda-Arizmendi, Diana Fimbres-Olivarria, Anselmo Miranda-Baeza, Agustín Rascón-Chu, Jorge Marquez-Escalante, Jaime Lizardi-Mendoza, Mayra A. Méndez-Encinas, Elizabeth Carvajal-Millan. Sulfated polysaccharides from marine diatoms: Insight into molecular characteristics and biological activity[J]. AIMS Bioengineering, 2024, 11(1): 110-129. doi: 10.3934/bioeng.2024007







 DownLoad:
DownLoad: