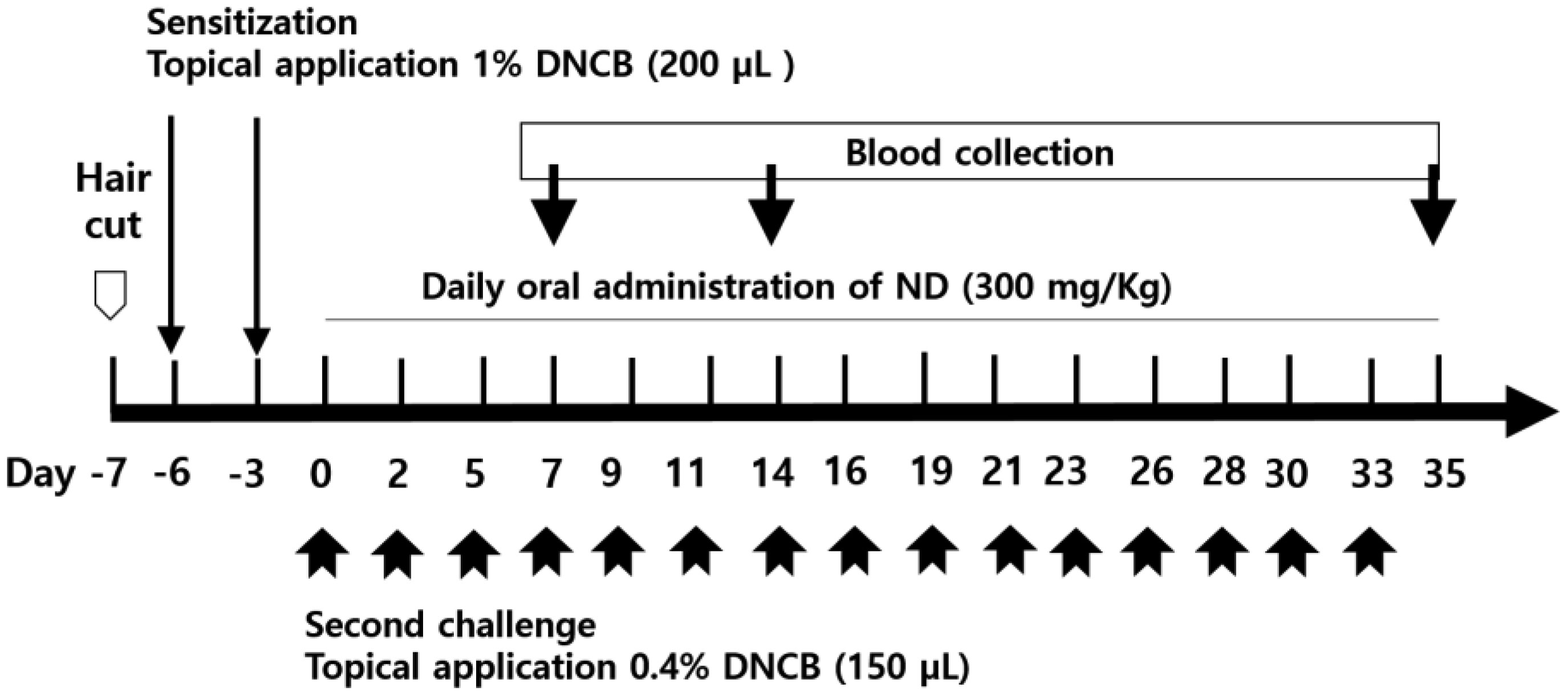
Atopic dermatitis (AD) is a skin disease. In this study, we observed the effects of Nandina domestica (ND) on AD-induced NC/Nga mice with 1-chloro-2,4-dinitrobenzene (DNCB)-treatment. ND significantly ameliorated the AD-skin signs with reducing of skin thickness and mast cell infiltration in the injuried skin of DNCB-induced AD model. Furthermore, ND reduced serum immunoglobulin E (IgE) levels and restored into normal condition the total cell number (TCN) in dorsal skin tissue, the axillary lymph node (ALN), and spleen after DNCB exposure. ND also decreased the number of CD23+/B220+ cells in the ALN and CD3+ cells in dorsal skin. Moreover, it decreased the interleukin (IL)-4, (IL)-5 and IL-13 levels but increased the interferon-γ levels in splenocytes. Immunohistofluorescence staining showed that ND significantly increased claudin1 and Sirt1 protein expressions in the AD-induced mice. Our results suggested that ND significantly ameliorates DCNB-induced AD.
Citation: Mi-Young Yun, Kyoung-Sook Park, Kyung-Ah Yoon, Hwa-Jung Choi. Leaf extracts of Nandina domestica Thunb ameliorate atopic dermatitis in NC/Nga mice treated with 1-chloro-2,4-dinitrobenzene (DNCB)[J]. AIMS Bioengineering, 2021, 8(2): 154-164. doi: 10.3934/bioeng.2021014
Atopic dermatitis (AD) is a skin disease. In this study, we observed the effects of Nandina domestica (ND) on AD-induced NC/Nga mice with 1-chloro-2,4-dinitrobenzene (DNCB)-treatment. ND significantly ameliorated the AD-skin signs with reducing of skin thickness and mast cell infiltration in the injuried skin of DNCB-induced AD model. Furthermore, ND reduced serum immunoglobulin E (IgE) levels and restored into normal condition the total cell number (TCN) in dorsal skin tissue, the axillary lymph node (ALN), and spleen after DNCB exposure. ND also decreased the number of CD23+/B220+ cells in the ALN and CD3+ cells in dorsal skin. Moreover, it decreased the interleukin (IL)-4, (IL)-5 and IL-13 levels but increased the interferon-γ levels in splenocytes. Immunohistofluorescence staining showed that ND significantly increased claudin1 and Sirt1 protein expressions in the AD-induced mice. Our results suggested that ND significantly ameliorates DCNB-induced AD.

| [1] |
Misery L, Huet F, Gouin O, et al. (2019) Current pharmaceutical developments in atopic dermatitis. Curr Opin Pharmacol 46: 7-13. doi: 10.1016/j.coph.2018.12.003

|
| [2] |
Brunner PM (2019) Early immunologic changes during the onset of atopic dermatitis. Ann Allergy Asthma Immunol 123: 152-157. doi: 10.1016/j.anai.2019.03.033
|
| [3] |
Bieber T (2008) Atopic dermatitis. N Engl J Med 358: 1483-1494. doi: 10.1056/NEJMra074081
|
| [4] |
Ponyai G, Hidvegi B, Nemeth I, et al. (2008) Contact and aeroallergens in adulthood atopic dermatitis. J Eur Acad Dermatol Venereol 22: 1346-1355. doi: 10.1111/j.1468-3083.2008.02886.x
|
| [5] |
Hamid Q, Boguniewicz M, Leung DY (1994) Differential in situ cytokine gene expression in acute versus chronic atopic dermatitis. J Clin Invest 94: 870-876. doi: 10.1172/JCI117408
|
| [6] |
Leung DY (2000) Atopic dermatitis: New insights and opportunities for therapeutic intervention. J Allergy Clin Immunol 105: 860-876. doi: 10.1067/mai.2000.106484
|
| [7] |
Elias PM, Hatano Y, Williams ML (2008) Basis for the barrier abnormality in atopic dermatitis: Outside-inside-outside pathogenic mechanisms. J Allergy Clin Immunol 121: 1337-1343. doi: 10.1016/j.jaci.2008.01.022
|
| [8] |
Boguniewicz M, Leung DYM (2011) Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol Rev 242: 233-246. doi: 10.1111/j.1600-065X.2011.01027.x
|
| [9] |
Shin KO, Ha DH, Kim JO, et al. (2020) Exosomes from human adipose tissue-derived mesenchymal stem cells promote epidermal barrier repair by inducing de novo synthesis of ceramides in atopic dermatitis. Cells 9: 680. doi: 10.3390/cells9030680
|
| [10] |
Schakel K, Dobel T, Bosselmann I (2014) Future treatment options for atopic dermatitis—small molecules and beyond. J Dermatol Sci 73: 91-100. doi: 10.1016/j.jdermsci.2013.11.009
|
| [11] |
Boguniewicz M, Alexis AF, Beck LA, et al. (2017) Expert perspectives on management of moderate-to-severe atopic dermatitis: A multidisciplinary consensus addressing current and emerging therapies. J Allergy Clin Immunol Pract 5: 1519-1531. doi: 10.1016/j.jaip.2017.08.005
|
| [12] | Akhtar N, Verma A, Pathak K (2017) Exploring preclinical and clinical effectiveness of nanoformulations in the treatment of atopic dermatitis: Safety aspects and patent reviews. Bull Fac Pharm Cairo Univ 55: 1-10. |
| [13] | Guo ZY, Zhang ZY, Xiao JQ, et al. (2018) Antibacterial effects of leaf extract of Nandina domestica and the underlined nechanism. Evid Based Complement Alternat Med 2018: 8298151. |
| [14] |
Bajpai VK, Yoon JI, Kang SC (2009) Antifungal potential of essential oil and various organic extracts of Nandina domestica Thunb. against skin infectious fungal pathogens. Appl Microbiol Biotechnol 83: 1127-1133. doi: 10.1007/s00253-009-2017-5
|
| [15] | Yun MY, Zhe W, Choi HJ (2020) Anti-inflammatory effect of Nandina domestica Thunb extracts possessing anti-oxidant property by regulating MAPKs signaling in LPS-induced inflammatory cells. J Altern Complement Integr Med 6: 114. |
| [16] | Guo ZY, Zhang ZY, Xiao JQ, et al. (2018) Antibacterial effects of leaf extract of Nandina domestica and the underlined mechanism. Evid Based Complement Alternat Med 2018: 8298151. |
| [17] |
Cabanillas B, Brehler AC, Novak N (2017) Atopic dermatitis phenotypes and the need for personalized medicin. Curr Opin Allergy Clin Immunol 17: 309-315. doi: 10.1097/ACI.0000000000000376
|
| [18] |
Akdis CA, Blaser K, Akdis M (2004) Apoptosis in tissue inflammation and allergic disease. Curr Opin Immunol 16: 717-723. doi: 10.1016/j.coi.2004.09.004
|
| [19] |
Verhagen J, Akdis M, Traidl-Hoffmann C, et al. (2006) Absence of T-regulatory cell expression and function in atopic dermatitis skin. J Allergy Clin Immunol 117: 176-183. doi: 10.1016/j.jaci.2005.10.040
|
| [20] |
Spergel JM, Mizoguchi E, Oettgen H, et al. (1999) Roles of TH1 and TH2 cytokines in a murine model of allergic dermatitis. J Clin Invest 103: 1103-1111. doi: 10.1172/JCI5669
|
| [21] |
Yoshimoto T, Yasuda K, Tanaka H, et al. (2009) Basophils contribute to TH2-IgE responses in vivo via IL-4 production and presentation of peptide–MHC class II complexes to CD4+ T cells. Nat Immunol 10: 706-712. doi: 10.1038/ni.1737
|
| [22] |
Leung DY, Bieber T (2003) Atopic dermatitis. Lancet 361: 151-160. doi: 10.1016/S0140-6736(03)12193-9
|
| [23] |
Liu FT, Goodarzi H, Chen HY (2011) IgE, mast cells, and eosinophils in atopic dermatitis. Clin Rev Allergy Immunol 41: 298-310. doi: 10.1007/s12016-011-8252-4
|
| [24] |
Ellingsen AR, Sorensen FB, Larsen JO, et al. (2001) Stereological quantification of lymphocytes in skin biopsies from atopic dermatitis patients. Acta Derm Venereol 81: 258-262. doi: 10.1080/00015550152572886
|
| [25] |
Parel Y, Chizzolini C (2004) CD4+ CD8+ double positive (DP) T cells in health and disease. Autoimmun Rev 3: 215-220. doi: 10.1016/j.autrev.2003.09.001
|
| [26] |
Tsukita S, Furuse M, Itoh M (2001) Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol 2: 285-293. doi: 10.1038/35067088
|
| [27] |
Brandner JM, Kief S, Grund C, et al. (2002) Organization and formation of the tight junction system in human epidermis and cultured keratinocytes. Eur J Cell Biol 81: 253-263. doi: 10.1078/0171-9335-00244
|
| [28] |
Haigis MC, Guarente LP (2006) Mammalian sirtuins—emerging roles in physiology, aging, and calorie restriction. Gene Dev 20: 2913-2921. doi: 10.1101/gad.1467506
|
| [29] |
Ming M, Shea CR, Guo X, et al. (2010) Regulation of global genome nucleotide excision repair by SIRT1 through xeroderma pigmentosum C. Proc Natl Acad Sci USA 107: 22623-22628. doi: 10.1073/pnas.1010377108
|



Figures(4) / Tables(1)

Mi-Young Yun, Kyoung-Sook Park, Kyung-Ah Yoon, Hwa-Jung Choi. Leaf extracts of Nandina domestica Thunb ameliorate atopic dermatitis in NC/Nga mice treated with 1-chloro-2,4-dinitrobenzene (DNCB)[J]. AIMS Bioengineering, 2021, 8(2): 154-164. doi: 10.3934/bioeng.2021014







 DownLoad:
DownLoad: