Citation: Nelson Mpumi, Revocatus L. Machunda, Kelvin M. Mtei, Patrick A. Ndakidemi. Insecticidal Efficacy of Syzygium aromaticum, Tephrosia vogelii and Croton dichogamus Extracts against Plutella xylostella and Trichoplusia ni on Brassica oleracea crop in Northern Tanzania[J]. AIMS Agriculture and Food, 2021, 6(1): 185-202. doi: 10.3934/agrfood.2021012

| [1] | Baidoo P, Mochiah M (2016) Comparing the effectiveness of garlic (Allium sativum L.) and hot pepper (Capsicum frutescens L.) in the management of the major pests of cabbage Brassica oleracea (L.). Sustainable Agric Res 5: 83-91. |
| [2] | Mpumi N, Machunda RS, Mtei KM, et al. (2020) Selected insect pests of economic importance to Brassica oleracea, their control strategies and the potential threat to environmental pollution in Africa. Sustainability 12: 1-22. |
| [3] | Mudzingwa S, Muzemu S, Chitamba J (2013) Pesticidal efficacy of crude aqueous extracts of Tephrosia vogelii L., Allium sativum L. and Solanum incanum L. in controlling aphids (Brevicoryne brassicae L.) in rape (Brassica napus L.). J Agric Res 2: 157-163. |
| [4] | Norman C, Shealy M (2007) Illustrated Encyclopedia of Healing Remedies. Toledo, OH, USA: Elements Book Inc. |
| [5] | Gibson A (2012) Colewart and the cole crops. Los Angeles: University of California Los Angeles. |
| [6] | Baidoo P, Adam J (2012) The Effects of Extracts of Lantana camara (L.) and Azadirachta indica (A. Juss) on the Population Dynamics of Plutella xylostella, Brevicoryne brassicae and Hellula undalis on Cabbage. Sustainable Agric Res 1: 229-234. |
| [7] | Furlong MJ, Wright DJ, Dosdall LM (2013) Diamondback moth ecology and management: problems, progress, and prospects. Annu Rev Entomol 58: 517-541. |
| [8] | Labou B, Brévault T, Sylla S, et al. (2017) Spatial and temporal incidence of insect pests in farmers' cabbage fields in Senegal. Int J Trop Insect Sci 37: 225-233. |
| [9] | Begna F, Damtew T (2015) Evaluation of four botanical insecticides against Diamondback Moth, Plutella Xylostella L.(Lepidoptera: Plutellidae) on head cabbage in the central rift valley of Ethiopia. Sky J Agric Res 4: 97-105. |
| [10] | Timbilla J, Nyarko K (2004) A survey of cabbage production and constraints in Ghana. Ghana J Agric Sci 37: 93-101. |
| [11] | Fening K, Amoabeng B, Adama I, et al. (2013) Sustainable management of two key pests of cabbage, Brassica oleracea var. capitata L.(Brassicaceae), using homemade extracts from garlic and hot pepper. Org Agric 3: 163-173. |
| [12] | Capinera J (2001) Handbook of vegetable pests. Eds, editor: Cambridge, MA, USA: Academic Press. |
| [13] | Mpumi N, Mtei K, Machunda R, et al. (2016) The toxicity, persistence and mode of actions of selected botanical pesticides in Africa against insect pests in common beans, P. vulgaris: a review. Am J Plant Sci 7: 138-151. |
| [14] | Labou B, Brévault T, Bordat D, et al. (2016) Determinants of parasitoid assemblages of the diamondback moth, Plutella xylostella, in cabbage farmer fields in Senegal. Crop Prot 89: 6-11. |
| [15] | Mkenda P, Mwanauta R, Stevenson PC, et al. (2015) Extracts from field margin weeds provide economically viable and environmentally benign pest control compared to synthetic pesticides. PLoS ONE 10: 1-14. |
| [16] | Mkindi A, Mpumi N, Tembo Y, et al. (2017) Invasive weeds with pesticidal properties as potential new crops. Ind Crops Prod 110: 113-122. |
| [17] | Ondieki JJ (1996) The current state of pesticide management in Sub-Saharan Africa. Sci Total Environ 188: 30-34. |
| [18] | Weinberger K, Srinivasan R (2009) Farmers' management of cabbage and cauliflower pests in India and their approaches to crop protection. J Asia-Pac Entomol 12: 253-259. |
| [19] | Amoabeng BW, Gurr GM, Gitau CW, et al. (2014) Cost: Benefit analysis of botanical insecticide use in cabbage: Implications for smallholder farmers in developing countries. Crop Prot 57: 71-76. |
| [20] | de Cássia Seffrin R, Shikano I, Akhtar Y, et al. (2010) Effects of crude seed extracts of Annona atemoya and Annona squamosa L. against the cabbage looper, Trichoplusia ni in the laboratory and greenhouse. Crop Prot 29: 20-24. |
| [21] | Mochiah M, Banful B, Fening K, et al. (2011) Botanicals for the management of insect pests in organic vegetable production. J Entomol Nematol 3: 85-97. |
| [22] | Isman MB (2006) Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu Rev Entomol 51: 45-66. |
| [23] | Orwa C, Mutua A, Kindt R, et al. (2009) Agroforestree database: a tree species reference and selection guide version 4.0. World Agroforestry Centre ICRAF, Nairobi, KE. |
| [24] | Munthali MG, Gachene CK, Sileshi GW, et al. (2014) Amendment of Tephrosia Improved Fallows with Inorganic Fertilizers Improves Soil Chemical Properties, N Uptake, and Maize Yield in Malawi. Int J Agron 2014: 1-9. |
| [25] | Kamatou GP, Vermaak I, Viljoen AM (2012) Eugenol from the remote Maluku Islands to the international market place: A review of a remarkable and versatile molecule. Molecules 17: 6953-6981. |
| [26] | Tian BL, Liu QZ, Liu ZL, et al. (2015) Insecticidal potential of clove essential oil and its constituents on Cacopsylla chinensis (Hemiptera: Psyllidae) in laboratory and field. J Econ Entomol 108: 957-961. |
| [27] | Araujo AFdO, Ribeiro-Paes JT, Deus JTd, et al. (2016) Larvicidal activity of Syzygium aromaticum (L.) Merr and Citrus sinensis (L.) Osbeck essential oils and their antagonistic effects with temephos in resistant populations of Aedes aegypti. Memórias do Instituto Oswaldo Cruz 111: 443-449. |
| [28] | Silva CG, Oliveira J, da Camara CA (2018) Insecticidal activity of the ethanolic extract from Croton species against Plutella xylostella L.(Lepidoptera: Plutellidae). Revista Facultad Nacional de Agronomía Medellín 71: 8543-8551. |
| [29] | Aldhaher A, Langat M, Ndunda B, et al. (2017) Diterpenoids from the roots of Croton dichogamus Pax. Phytochemistry 144: 1-8. |
| [30] | Belmain SR, Amoah BA, Nyirenda SP, et al. (2012) Highly variable insect control efficacy of Tephrosia vogelii chemotypes. J Agric Food Chem 60: 10055-10063. |
| [31] | Patra S, Dhote V, Alam S, et al. (2013) Population dynamics of major insect pests and their natural enemies on cabbage under new alluvial zone of West Bengal. J Plant Prot Sci 5: 42-49. |
| [32] | Tanyi CB, Ngosong C, Ntonifor NN (2018) Effects of climate variability on insect pests of cabbage: adapting alternative planting dates and cropping pattern as control measures. Chem Biol Technol Agric 5: 1-11. |
| [33] | Ayalew G, Ogol C (2006) Occurrence of the diamondback moth (Plutella xylostella L.) and its parasitoids in Ethiopia: influence of geographical region and agronomic traits. J Appl Entomol 130: 343-348. |
| [34] | Grzywacz D, Rossbach A, Rauf A, et al. (2010) Current control methods for diamondback moth and other brassica insect pests and the prospects for improved management with lepidopteran-resistant Bt vegetable brassicas in Asia and Africa. Crop Prot 29: 68-79. |
| [35] | Belmain S, Haggar J, Holt J, et al. (2013) Managing legume pests in sub-Saharan Africa: Challenges and prospects for improving food security and nutrition through agro-ecological intensification. Chatham Maritime (United Kingdom): Natural Resources Institute, University of Greenwich, 34. |
| [36] | Belmain S, Stevenson P (2001) Ethnobotanicals in Ghana: Reviving and modernising age-old farmer practice. Pestic Outlook 12: 233-238. |
| [37] | Do Ngoc Dai TDT, Ogunmoye A, Eresanya OI, et al. (2015) Chemical constituents of essential oils from the leaves of Tithonia diversifolia, Houttuynia cordata and Asarum glabrum grown in Vietnam. Am J Essent Oils Nat Prod 2: 17-21. |
| [38] | Grzywacz D, Stevenson P, Mushobozi WL, et al. (2014) The use of indigenous ecological resources for pest control in Africa. Food Secur 6: 71-86. |
| [39] | Ileke K, Oni M (2011) Toxicity of some plant powders to maize weevil, Sitophilus zeamais (motschulsky)[Coleoptera: Curculiondae] on stored wheat grains (Triticum aestivum). Afr J Biotechnol 6: 3043-3048. |
| [40] | Kamanula J, Sileshi GW, Belmain SR, et al. (2010) Farmers' insect pest management practices and pesticidal plant use in the protection of stored maize and beans in Southern Africa. Int J Pest Manag 57: 41-49. |
| [41] | Mkenda PA, Mtei K, Ndakidemi PA (2014) Pesticidal efficacy of Tephrosia vogelii and Tithonia diversifolia against field insect pests of common beans[Phaseolus vulgaris L.]. within African farming communities. Afr J Appl Agric Sci Technol 2: 9-26. |
| [42] | Mwanauta RW, Mtei K, Ndakidemi PA (2014) Prospective Bioactive Compounds from Vernonia amygdalina, Lippia javanica, Dysphania ambrosioides and Tithonia diversifolia in Controlling Legume Insect Pests. Agric Sci 5: 1129-1139. |
| [43] | Khater HF (2012) Prospects of botanical biopesticides in insect pest management. Pharmacologia 3: 641-656. |
| [44] | Marchioro C, Foerster L (2011) Development and survival of the diamondback moth, Plutella xylostella (L.)(Lepidoptera: Yponomeutidae) as a function of temperature: effect on the number of generations in tropical and subtropical regions. Neotrop Entomol 40: 533-541. |
| [45] | Kobori Y, Amano H (2003) Effect of rainfall on a population of the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Appl Entomol Zool 38: 249-253. |
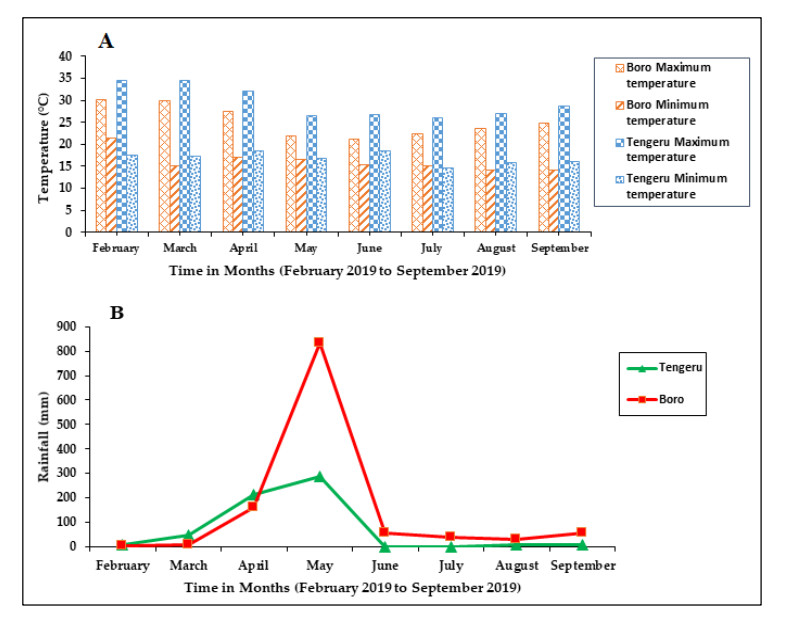
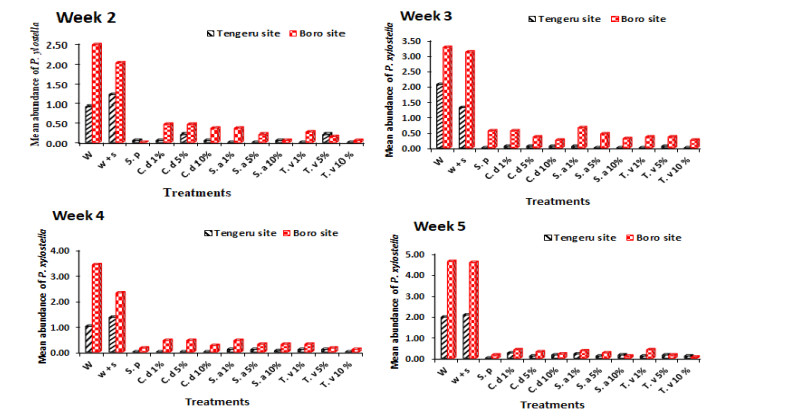
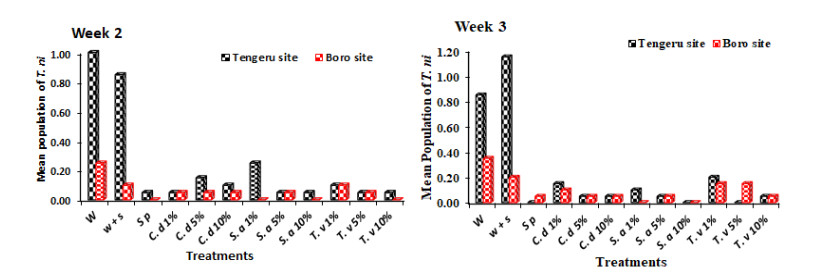
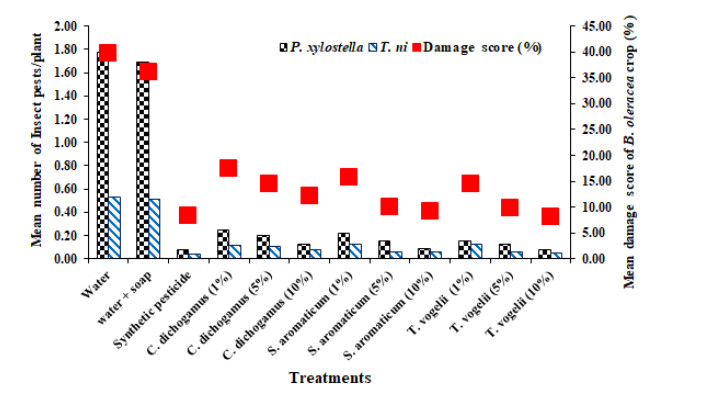
Figures(5) / Tables(4)

Nelson Mpumi, Revocatus L. Machunda, Kelvin M. Mtei, Patrick A. Ndakidemi. Insecticidal Efficacy of Syzygium aromaticum, Tephrosia vogelii and Croton dichogamus Extracts against Plutella xylostella and Trichoplusia ni on Brassica oleracea crop in Northern Tanzania[J]. AIMS Agriculture and Food, 2021, 6(1): 185-202. doi: 10.3934/agrfood.2021012







 DownLoad:
DownLoad: