Citation: Baldi Elena, Marino Grazia, Muzzi Enrico, Marzadori Claudio, Ciavatta Claudio, Tavoni Marta, Di Giosia Matteo, Calvaresi Matteo, Falini Giuseppe, Zerbetto Francesco, Toselli Moreno. Effectiveness of synthetic calcite doped with Fe-EDDHSA as a slow-release Fe source: In-vitro experiment on kiwifruit (Actinidia chinensis var. deliciosa) plants[J]. AIMS Agriculture and Food, 2019, 4(1): 127-135. doi: 10.3934/agrfood.2019.1.127

| [1] |
Tagliavini M, Rombolà AD (2001) Iron deficiency and chlorosis in orchard and vineyard ecosystems. Eur J Agron 15: 71–92. doi: 10.1016/S1161-0301(01)00125-3

|
| [2] | Lindsay WL (1979) Chemical equilibria in soils. John Wiley and Sons Inc., NY, USA. |
| [3] |
Marschner H, Römheld V (1994) Strategies of plants for acquisition of iron. Plant Soil 165: 261–274. doi: 10.1007/BF00008069
|
| [4] | Romheld V, Marschner H (1986) Mobilization of iron in the rhizosphere of different plant species. Adv Plant Nutr 2: 155–204. |
| [5] |
Rombolà AD, Brüggemann W, López-Millán AF, et al. (2002) Biochemical responses to iron deficiency in kiwifruit (Actinidia deliciosa). Tree Physiol 22: 869–875. doi: 10.1093/treephys/22.12.869
|
| [6] |
Farokhzad OC, Langer R (2009) Impact of nanotechnology on drug delivery. ACS Nano 3: 16–20. doi: 10.1021/nn900002m
|
| [7] | Sun T, Zhang YS, Pang B, et al. (2014) Engineered nanoparticles for drug delivery in cancer therapy. Angew Chem Int Ed 53: 12320–12364. |
| [8] | Jampílek J, Kráľová K (2017) Nanomaterials for delivery of nutrients and growth-promoting compounds to plants. Nanotechnology, 177–226. |
| [9] |
Marino G, Marcolini G, Toselli M (2006) Evaluation of the activity of aqueous extracts of some organic waste materials on in vitro-cultured shoots of 'M9' apple rootstock. J Hortic Sci Biotechnol 81: 1015–1020. doi: 10.1080/14620316.2006.11512164
|
| [10] |
Capuana M (2011) Heavy metals and woody plants-biotechnologies for phytoremediation. iForest-Biogeosciences and Forestry 4: 7. doi: 10.3832/ifor0555-004
|
| [11] |
Lombardi L, Sebastiani L, Vitagliano C (2003) Physiological, biochemical, and molecular effects of in vitro induced iron deficiency in peach rootstock Mr.S2/5. J Plant Nutr 26: 2149–2163. doi: 10.1081/PLN-120024271
|
| [12] |
Karam NS, Ereifej KI, Shibli RA, et al. (1998) Metal concentrations, growth, and yield of potato produced from in-vitro plantlets or microtubers and grown in municipal solid-waste-amended substrates. J Plant Nutr 21: 725–739. doi: 10.1080/01904169809365438
|
| [13] |
Houshmand S, Arzani A, Maibody SAM, et al (2005) Evaluation of salt-tolerant genotypes of durum wheat derived from in vitro and field experiments. Field Crop Res 91: 345–354. doi: 10.1016/j.fcr.2004.08.004
|
| [14] |
Marino G, Beghelli S, Rombolà AD, et al. (2000) In vitro performance at high culture pH and in vivo responses to Fe-deficiency of leaf-derived quince BA 29 (Cydonia oblonga) somaclones regenerated at variable medium pH. J Hortic Sci Biotechnol 75: 433–440. doi: 10.1080/14620316.2000.11511264
|
| [15] | Bunnag S, Dolcet-Sanjuan R, Mok DWS, et al. (1996) Responses of two somaclonal variants of quince (Cydonia oblonga) to iron deficiency in the greenhouse and field. J Am Soc Hortic Sci 12: 1054–1058. |
| [16] | Marino G, Bertazza G, Buscaroli C (1998) In vivo growth and tolerance to lime-induced iron chlorosis of leaf-derived cvs. Tomuri and Hayward kiwifruit (Actinidia deliciosa) somaclones. J Hortic Sci Biotechnol 73: 670–675. |
| [17] |
Marino G, Bertazza G (1998) Selection-pressure effects of medium pH during regeneration on successive performance of leaf-derived 'Tomuri' and 'Hayward' kiwifruit (Actinidia deliciosa) somaclones cultured on proliferation culture media with variable pH. J Hortic Sci Biotechnol 73: 664–669. doi: 10.1080/14620316.1998.11511030
|
| [18] |
Baldi E, Marino G, Toselli M, et al. (2018) Delivery systems for agriculture: Fe-EDDHSA/CaCO3 hybrid crystals as adjuvants for prevention of iron chlorosis. Chem Commun 54: 1635–1638. doi: 10.1039/C7CC08215K
|
| [19] |
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15: 473–497. doi: 10.1111/j.1399-3054.1962.tb08052.x
|
| [20] |
Magnabosco G, Giosia MD, Polishchuk I, et al. (2015) Calcite single crystals as hosts for atomic-scale entrapment and slow release of drugs. Adv Health Mater 4: 1510–1516. doi: 10.1002/adhm.201500170
|
| [21] | Kingston HM (1988) Microwave assisted acid digestion of siliceous and organically-based matrices, Method 3052. U.S. Environmental Protection Agency IAG DWI-393254-01-0, Quarterly Report, January 1–March 31. |
| [22] | George EF (1993) Plant Propagation by tissue culture, Part 1, The technology. Edington, Wilts: Exegetics Ltd, 573. |
| [23] | Lee ECM, DeFossard RA (1997) Some factors affecting multiple bud formation of strawberry (Fragaria ananassa Duchesne) in vitro. Acta Hort 78: 187–195. |
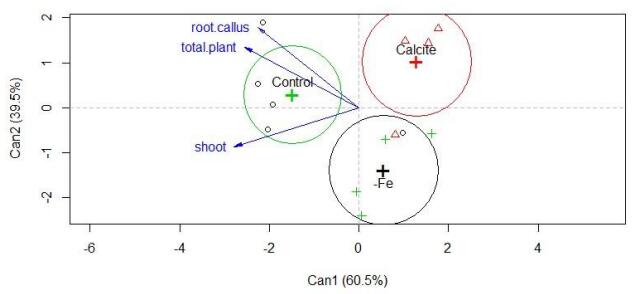
Figures(1) / Tables(3)

Baldi Elena, Marino Grazia, Muzzi Enrico, Marzadori Claudio, Ciavatta Claudio, Tavoni Marta, Di Giosia Matteo, Calvaresi Matteo, Falini Giuseppe, Zerbetto Francesco, Toselli Moreno. Effectiveness of synthetic calcite doped with Fe-EDDHSA as a slow-release Fe source: In-vitro experiment on kiwifruit (Actinidia chinensis var. deliciosa) plants[J]. AIMS Agriculture and Food, 2019, 4(1): 127-135. doi: 10.3934/agrfood.2019.1.127







 DownLoad:
DownLoad: