Previous studies have shown controversial results regarding the pro- or anticonvulsant effects of tramadol. Additionally, the underlying mechanism of seizure induction or alleviation by tramadol has not been fully understood. In the current study, the effects of tramadol on pentylenetetrazole (PTZ)-induced seizure and the possible involvement of the N-methyl-D-aspartate (NMDA) pathway were assessed in mice.
Male Naval Medical Research Institute (NMRI) mice were treated with intravenous infusion of PTZ in order to induce clonic seizures and determine seizure threshold. Tramadol was injected intraperitoneally (0.1–150 mg/kg) 30 minutes prior to elicitation of seizures. The possible effects of intraperitoneal injections of NMDA receptor antagonists, ketamine (0.5 mg/kg) and MK-801 (0.5 mg/kg) on the anticonvulsant property of tramadol were investigated subsequently.
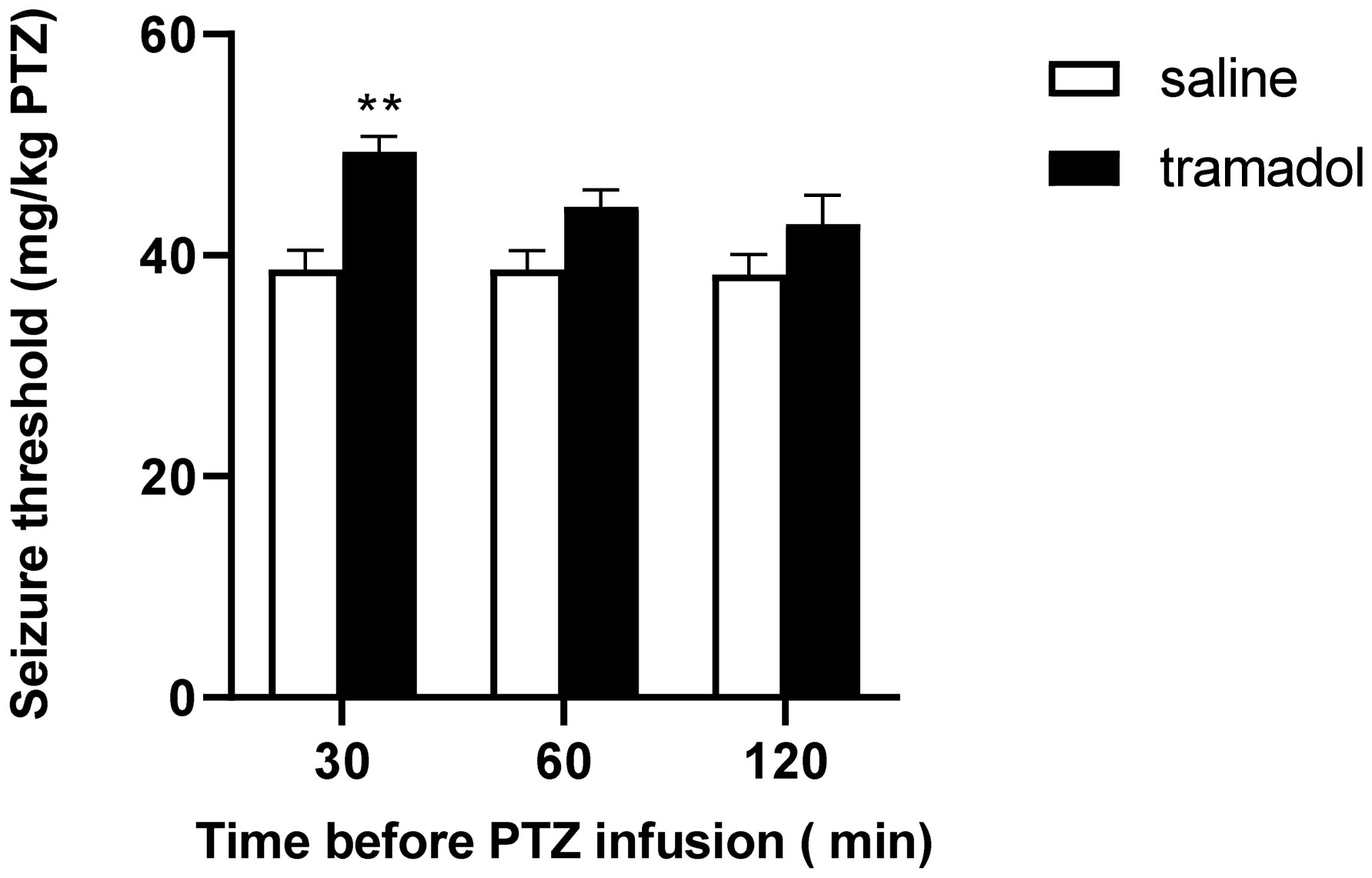
Tramadol (1–100 mg/kg) increased PTZ-induced seizure threshold in a dose-dependent, time-independent manner, with optimal anticonvulsant effect at a dose of 100 mg/kg. Acute administration of either ketamine (0.5 mg/kg) or MK-801 (0.5 mg/kg) potentiated the anticonvulsant effect of a subeffective dose of tramadol (0.3 mg/kg).
These results suggest a possible role of the NMDA pathway in the anticonvulsant effect of tramadol.
Citation: Mazyar Zahir, Amir Rashidian, Mohsen Hoseini, Reyhaneh Akbarian, Mohsen Chamanara. Pharmacological evidence for the possible involvement of the NMDA receptor pathway in the anticonvulsant effect of tramadol in mice[J]. AIMS Neuroscience, 2022, 9(4): 444-453. doi: 10.3934/Neuroscience.2022024
Previous studies have shown controversial results regarding the pro- or anticonvulsant effects of tramadol. Additionally, the underlying mechanism of seizure induction or alleviation by tramadol has not been fully understood. In the current study, the effects of tramadol on pentylenetetrazole (PTZ)-induced seizure and the possible involvement of the N-methyl-D-aspartate (NMDA) pathway were assessed in mice.
Male Naval Medical Research Institute (NMRI) mice were treated with intravenous infusion of PTZ in order to induce clonic seizures and determine seizure threshold. Tramadol was injected intraperitoneally (0.1–150 mg/kg) 30 minutes prior to elicitation of seizures. The possible effects of intraperitoneal injections of NMDA receptor antagonists, ketamine (0.5 mg/kg) and MK-801 (0.5 mg/kg) on the anticonvulsant property of tramadol were investigated subsequently.
Tramadol (1–100 mg/kg) increased PTZ-induced seizure threshold in a dose-dependent, time-independent manner, with optimal anticonvulsant effect at a dose of 100 mg/kg. Acute administration of either ketamine (0.5 mg/kg) or MK-801 (0.5 mg/kg) potentiated the anticonvulsant effect of a subeffective dose of tramadol (0.3 mg/kg).
These results suggest a possible role of the NMDA pathway in the anticonvulsant effect of tramadol.

| [1] |
Dayer P, Collart L, Desmeules J (1994) The pharmacology of tramadol. Drugs 47: 3-7. https://doi.org/10.2165/00003495-199400471-00003 
|
| [2] |
Roca-Vinardell A, Berrocosobcd E, Llorca-Torralbaacd M, et al. (2018) Involvement of 5-HT(1A/1B) receptors in the antinociceptive effect of paracetamol in the rat formalin test. Neurobiol Pain 3: 15-21. https://doi.org/10.1016/j.ynpai.2018.01.004
|
| [3] |
Ogata J, Minami K, Uezono Y, et al. (2004) The inhibitory effects of tramadol on 5-hydroxytryptamine type 2C receptors expressed in Xenopus oocytes. Anesth Analg 98: 1401-1406. https://doi.org/10.1213/01.ANE.0000108963.77623.A4
|
| [4] |
Kahn LH, Alderfer RJ, Graham DJ (1997) Seizures reported with tramadol. Jama 278: 1661. https://doi.org/10.1001/jama.1997.03550200037026
|
| [5] |
Potschka H, Friderichs E, Löscher W (2000) Anticonvulsant and proconvulsant effects of tramadol, its enantiomers and its M1 metabolite in the rat kindling model of epilepsy. Br J Pharmacol 131: 203-12. https://doi.org/10.1038/sj.bjp.0703562
|
| [6] |
Lesani A., et al. (2010) Involvement of the nitric oxide pathway in the anticonvulsant effect of tramadol on pentylenetetrazole-induced seizures in mice. Epilepsy Behav 19: 290-5. https://doi.org/10.1016/j.yebeh.2010.08.006
|
| [7] |
Manocha A, Sharma KK, Mediratta PK (2005) On the mechanism of anticonvulsant effect of tramadol in mice. Pharmacol Biochem Behav 82: 74-81. https://doi.org/10.1016/j.pbb.2005.07.013
|
| [8] |
Avoli M, D'Antuonob M, Louvelc J, et al. (2002) Network and pharmacological mechanisms leading to epileptiform synchronization in the limbic system in vitro. Prog Neurobiol 68: 167-207. https://doi.org/10.1016/S0301-0082(02)00077-1
|
| [9] |
Morimoto K, Fahnestock M, Racine RJ (2004) Kindling and status epilepticus models of epilepsy: rewiring the brain. Prog Neurobiol 73: 1-60. https://doi.org/10.1016/j.pneurobio.2004.03.009
|
| [10] |
Furukawa H, Singh SK, Mancusso R, et al. (2005) Subunit arrangement and function in NMDA receptors. Nature 438: 185-92. https://doi.org/10.1038/nature04089
|
| [11] | Blanke M.L., VanDongen A.M.J. (2009) Frontiers in Neuroscience Activation Mechanisms of the NMDA Receptor, in Biology of the NMDA Receptor. Boca Raton (FL): CRC Press/Taylor & Francis Copyright © 2009, Taylor & Francis Group, LLC. |
| [12] |
Newcomer JW, Farber NB, Olney JW (2000) NMDA receptor function, memory, and brain aging. Dialogues Clin Neurosci 2: 219-32. https://doi.org/10.31887/DCNS.2000.2.3/jnewcomer
|
| [13] |
Adell A (2020) Brain NMDA Receptors in Schizophrenia and Depression. Biomolecules 10: 947. https://doi.org/10.3390/biom10060947
|
| [14] |
Haj-Mirzaian A, Amiri S, Amini-Khoei H, et al. (2018) Involvement of NO/NMDA-R pathway in the behavioral despair induced by amphetamine withdrawal. Brain Res Bull 139: 81-90. https://doi.org/10.1016/j.brainresbull.2018.02.001
|
| [15] |
Ostadhadi S, Norouzi-Javidan A, Chamanara M, et al. (2017) Involvement of NMDA receptors in the antidepressant-like effect of tramadol in the mouse forced swimming test. Brain Res Bull 134: 136-141. https://doi.org/10.1016/j.brainresbull.2017.07.016
|
| [16] |
Jafari-Sabet M, Mofidi H, Attarian-Khosroshahi MS (2018) NMDA receptors in the dorsal hippocampal area are involved in tramadol state-dependent memory of passive avoidance learning in mice. Can J Physiol Pharmacol 96: 45-50. https://doi.org/10.1139/cjpp-2017-0228
|
| [17] |
Zahir M, Shariatzadeh S, Khosravi A, et al. (2021) High risk of drug toxicity in social isolation stress due to liver dysfunction: Role of oxidative stress and inflammation. Brain Behav 11: e2317. https://doi.org/10.1002/brb3.2317
|
| [18] |
Amiri S, et al. (2014) Involvement of the nitrergic system in the proconvulsant effect of social isolation stress in male mice. Epilepsy Behav 41: 158-63. https://doi.org/10.1016/j.yebeh.2014.09.080
|
| [19] |
Bahremand A, Nasrabady SE, Ziai P, et al. (2010) Involvement of nitric oxide-cGMP pathway in the anticonvulsant effects of lithium chloride on PTZ-induced seizure in mice. Epilepsy Res 89: 295-302. https://doi.org/10.1016/j.eplepsyres.2010.02.001
|
| [20] | Manocha A, Sharma KK, Mediratta PK (2001) Possible mechanism of anticonvulsant effect of ketamine in mice. Indian J Exp Biol 39: 1002-8. |
| [21] |
Dowell D, Haegerich TM, Chou R (2016) CDC Guideline for Prescribing Opioids for Chronic Pain - United States, 2016. MMWR Recomm Rep 65: 1-49. https://doi.org/10.15585/mmwr.rr6501e1
|
| [22] |
Rehni AK, Singh I, Kumar M (2008) Tramadol-induced seizurogenic effect: a possible role of opioid-dependent gamma-aminobutyric acid inhibitory pathway. Basic Clin Pharmacol Toxicol 103: 262-6. https://doi.org/10.1111/j.1742-7843.2008.00276.x
|
| [23] |
Atici S, Cinel L, Cinel I, et al. (2004) Opioid neurotoxicity: comparison of morphine and tramadol in an experimental rat model. Int J Neurosci 114: 1001-11. https://doi.org/10.1080/00207450490461314
|
| [24] |
Mishra P, et al. (2019) Application of student's t-test, analysis of variance, and covariance. Ann Card Anaesth 22: 407-411. https://doi.org/10.4103/aca.ACA_94_19
|
| [25] |
Samadi M, Singh U, Pandey CM, et al. (2021) Caffeine attenuates seizure and brain mitochondrial disruption induced by Tramadol: the role of adenosinergic pathway. Drug Chem Toxicol 44: 613-619. https://doi.org/10.1080/01480545.2019.1643874
|
| [26] |
Hansen KB, Yi F, Perszyk RE, et al. (2017) NMDA Receptors in the Central Nervous System. Methods Mol Biol 1677: 1-80. https://doi.org/10.1007/978-1-4939-7321-7_1
|
| [27] |
Witkin JM, Cerne R, Newman AH, et al. (2021) N-Substituted-3-alkoxy-derivatives of dextromethorphan are functional NMDA receptor antagonists in vivo: Evidence from an NMDA-induced seizure model in rats. Pharmacol Biochem Behav 203: 173154. https://doi.org/10.1016/j.pbb.2021.173154
|
| [28] |
Nasiri-Boroujeni S, Rahimi-Madiseh M, Lorigooini Z, et al. (2021) NMDA Receptor Mediates the Anticonvulsant Effect of Hydroalcoholic Extract of Artemisia persica in PTZ-Induced Seizure in Mice. Evid Based Complement Alternat Med 2021: 6422451. https://doi.org/10.1155/2021/6422451
|
| [29] | Roodsari SK, Bahramnejad E, Rahimi N, et al. (2019) Methadone's effects on pentylenetetrazole-induced seizure threshold in mice: NMDA/opioid receptors and nitric oxide signaling. Ann N Y Acad Sci 1449: 25-35. https://doi.org/10.1111/nyas.14043 |
| [30] |
Jafari-Sabet M, Amiri S, Ataee R (2018) Cross state-dependency of learning between tramadol and MK-801 in the mouse dorsal hippocampus: involvement of nitric oxide (NO) signaling pathway. Psychopharmacology (Berl) 235: 1987-1999. https://doi.org/10.1007/s00213-018-4897-5
|



Figures(4)

Mazyar Zahir, Amir Rashidian, Mohsen Hoseini, Reyhaneh Akbarian, Mohsen Chamanara. Pharmacological evidence for the possible involvement of the NMDA receptor pathway in the anticonvulsant effect of tramadol in mice[J]. AIMS Neuroscience, 2022, 9(4): 444-453. doi: 10.3934/Neuroscience.2022024







 DownLoad:
DownLoad: