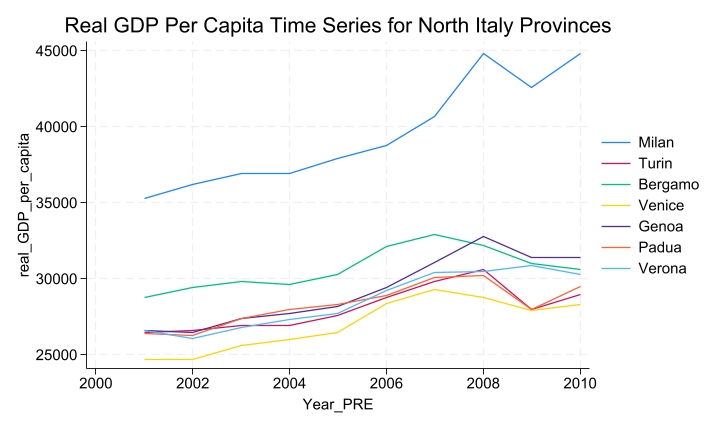
This paper aimed to investigate the economic impact of building a next-generation stadium (for example, Juventus Stadium or Allianz Stadium) on real gross domestic product (GDP) per capita in the city of Turin, using an econometric methodology known as the synthetic control method. The methodology compares the post-real GDP per capita trajectory of the treated provincial economy with that of a synthetic combination of similar, but untreated, provincial economies. The analysis showed that building a next-generation stadium had a short-term effect on real GDP per capita, with an increase of approximately 2% in the year of construction (2011). Additionally, the analysis showed a spring-back effect, where in the year following the stadium's construction, the real GDP per capita is slightly lower than what is projected by the synthetic control (around 0.85%). Moreover in the subsequent years, there seems to be a small positive structural effect of the treatment since the observed outcome is always higher than the synthetic outcome. Finally, the analysis also highlights an unexpected growth in real GDP per capita compared to the synthetic control, amounting for 0.5% in the year the stadium is announced (2008). Unlike prior studies, which have merely identified correlations, this research provides the first evidence of a causal relationship between the construction of a stadium and changes in the well-being of residents within the metropolitan area where the stadium is located.
Citation: Valerio Antolini. The economic impact of next-generation stadiums: evidence from the Juventus Stadium using synthetic control methodology[J]. National Accounting Review, 2024, 6(4): 531-547. doi: 10.3934/NAR.2024024
This paper aimed to investigate the economic impact of building a next-generation stadium (for example, Juventus Stadium or Allianz Stadium) on real gross domestic product (GDP) per capita in the city of Turin, using an econometric methodology known as the synthetic control method. The methodology compares the post-real GDP per capita trajectory of the treated provincial economy with that of a synthetic combination of similar, but untreated, provincial economies. The analysis showed that building a next-generation stadium had a short-term effect on real GDP per capita, with an increase of approximately 2% in the year of construction (2011). Additionally, the analysis showed a spring-back effect, where in the year following the stadium's construction, the real GDP per capita is slightly lower than what is projected by the synthetic control (around 0.85%). Moreover in the subsequent years, there seems to be a small positive structural effect of the treatment since the observed outcome is always higher than the synthetic outcome. Finally, the analysis also highlights an unexpected growth in real GDP per capita compared to the synthetic control, amounting for 0.5% in the year the stadium is announced (2008). Unlike prior studies, which have merely identified correlations, this research provides the first evidence of a causal relationship between the construction of a stadium and changes in the well-being of residents within the metropolitan area where the stadium is located.

| [1] |
Abadie A, Diamond A, Hainmueller J (2010) Synthetic control methods for comparative case studies: Estimating the effect of California's tobacco control program. J Am Stat Assoc 105: 493–505. https://doi.org/10.1198/jasa.2009.ap08746 doi: 10.1198/jasa.2009.ap08746

|
| [2] |
Abadie A, Gardeazabal J (2003) The economic costs of conflict: A case study of the Basque Country. Am Econ Rev 93: 113–132. https://doi.org/10.1257/000282803321455188 doi: 10.1257/000282803321455188
|
| [3] |
Arkhangelsky D, Athey S, Hirshberg DA, et al. (2021) Synthetic difference-in-differences. Am Econ Rev 111: 4088–4118. https://doi.org/10.1257/aer.20190159 doi: 10.1257/aer.20190159
|
| [4] |
Ashenfelter OC, Card D (1984) Using the longitudinal structure of earnings to estimate the effect of training programs. Rev Econ Stat 11: 648–660. https://doi.org/10.2307/1924810 doi: 10.2307/1924810
|
| [5] |
Baade RA, Dye RF (1998) Sports stadiums and area development: A critical review. Econ Dev Q 2: 265–275. https://doi.org/10.1177/089124248800200306 doi: 10.1177/089124248800200306
|
| [6] |
Baade RA, Tiehen LJ (1990) An analysis of major league baseball attendance, 1969–1987. J Sport Soc Issues 14: 265–275. https://doi.org/10.1177/019372359001400102 doi: 10.1177/019372359001400102
|
| [7] |
Baade RA (1996) Professional sports as catalysts for metropolitan economic development. J Urban Aff 18: 1–17. https://doi.org/10.1111/j.1467-9906.1996.tb00361.x doi: 10.1111/j.1467-9906.1996.tb00361.x
|
| [8] | Baade RA, Sanderson AR (1997) Cities under siege: how the changing financial structure of professional sports is putting cities at risk and what to do about it. |
| [9] | Bertrand M, Duflo E, Mullainathan S (1984) How much should we trust differences-in-differences estimates? Q J Econ 119: 249–275. https://doi.org/10.1162/003355304772839588 |
| [10] | Blanchard O (1993) Consumption and the Recession of 1990–1991. Am Econ Rev 83: 270–274. |
| [11] | Blomstrom M, Lipsey RE, Zejan M (1992) What explains developing country growth? Available from: https://www.nber.org/system/files/working_papers/w4132/w4132.pdf. |
| [12] | Carroll CD, Fuhrer JC, Wilcox DW (1994) Does consumer sentiment forecast household spending? If so, why?. Am Econ Rev 84: 1397–1408. |
| [13] | Card D, Krueger AB (1993) Minimum wages and employment: A case study of the fast food industry in New Jersey and Pennsylvania. Am Econ Rev 90: 1397–1420. |
| [14] | Coates D, Humphreys BR (2008) Do economists reach a conclusion on subsidies for sports franchises, stadiums, and mega-events? Econ J Watch 5: 294–315. |
| [15] |
Firpo S, Possebom V (2018) Synthetic control method: Inference, sensitivity analysis and confidence sets. J Causal Inference 6: 20160026. https://doi.org/10.1515/jci-2016-0026 doi: 10.1515/jci-2016-0026
|
| [16] |
Formánek T (2019) GDP per capita in selected EU countries: Economic growth factors and spatio-temporal interactions examined at the NUTS2 level. J Int Stud 12: 119–133. https://doi.org/10.14254/2071-8330.2019/12-1/8 doi: 10.14254/2071-8330.2019/12-1/8
|
| [17] | Gennaioli N, Ma Y, Shleifer A (2016) Expectations and investment. NBER Macroeconomics Annual 30: 379–431. |
| [18] |
Ilter C (2017) What economic and social factors affect GDP per capita? A study on 40 countries. J Global Strategic Manage 11: 51–62. https://doi.org/10.20460/JGSM.2018.252 doi: 10.20460/JGSM.2018.252
|
| [19] |
Kreif N, Grieve R, Hangartner D, et al. (2016) Examination of the synthetic control method for evaluating health policies with multiple treated units. Health Econ 25: 1514–1528. https://doi.org/10.1002/hec.3258 doi: 10.1002/hec.3258
|
| [20] |
Mello M (2024) A kick for the GDP: the effect of winning the FIFA World Cup. Am Econ Rev 86: 1313–1341. https://doi.org/10.1111/obes.12627 doi: 10.1111/obes.12627
|
| [21] |
Miller PA (2002) The economic impact of sports stadium construction: The case of the construction industry in St. Louis, MO. J Urban Aff 24: 159–173. https://doi.org/10.1111/1467-9906.00120 doi: 10.1111/1467-9906.00120
|
| [22] |
Santo C (2005) The economic impact of sports stadiums: Recasting the analysis in context. J Urban Aff 27: 177–192. https://doi.org/10.1111/j.0735-2166.2005.00231.x doi: 10.1111/j.0735-2166.2005.00231.x
|
| [23] | Wu J (2012) Cluster analysis and K-means clustering: an introduction, In: Advances in K-means Clustering, Springer Theses. Springer, Berlin, Heidelberg, 1–16. https://doi.org/10.1007/978-3-642-29807-3_1 |






Figures(7) / Tables(3)

Valerio Antolini. The economic impact of next-generation stadiums: evidence from the Juventus Stadium using synthetic control methodology[J]. National Accounting Review, 2024, 6(4): 531-547. doi: 10.3934/NAR.2024024







 DownLoad:
DownLoad: