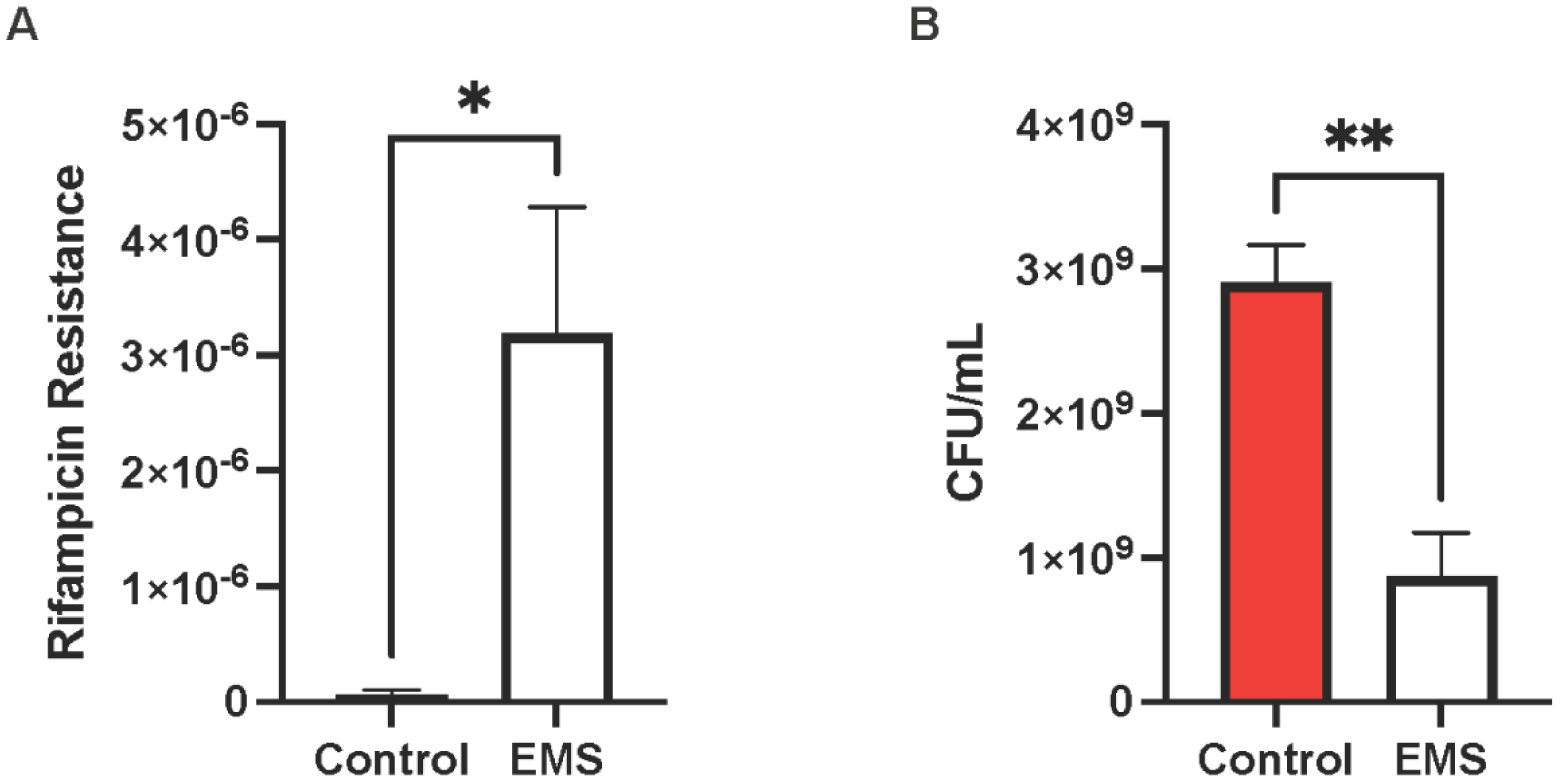
Staphylococcus lugdunensis is a coagulase-negative species responsible for a multitude of infections. These infections often resemble those caused by the more pathogenic staphylococcal species, Staphylococcus aureus, such as skin and soft tissue infections, prosthetic joint infections, and infective endocarditis. Despite a high mortality rate and infections that differ from other coagulase-negative species, little is known regarding S. lugdunensis pathogenesis. The objective of this study is to identify the essential factors for biofilm formation in S. lugdunensis. S. lugdunensis was mutagenized through ethyl methanesulfonate (EMS) exposure, and the individual cells were separated using a cell sorter and examined for biofilm formation at 8 hr and 24 hr timepoints. Mutations that resulted in either increased or decreased biofilm formation were sequenced to identify the genes responsible for the respective phenotypes. A mutation within the S. lugdunensis surface protein A (slsA) gene was common among all of the low biofilm formers, thus suggesting that high expression of this protein is important in biofilm formation. However, other mutations common among the mutants with decreased biofilm formation were in the putative divalent cation transport gene, mgtE. Conversely, a mutation in the gene that codes for the von Willebrand factor binding protein, vwbl, was common among the mutants with increased biofilm formation. Following proteinase K treatment, a significant dispersal of the S. lugdunensis biofilm matrix occurred, thus confirming the presence of primarily protein-mediated biofilms; this is in agreement with previous S. lugdunensis studies. Additionally, all low biofilm formers exhibited decreased protein levels (1.95–2.77 fold change) within the biofilm matrix, while no difference was observed with extracellular DNA (eDNA) or polysaccharides. This study presents a unique methodology to identify genes that affect biofilm formation and sheds light on S. lugdunensis pathogenesis.
Citation: McKenna J. Cruikshank, Justine M. Pitzer, Kimia Ameri, Caleb V. Rother, Kathryn Cooper, Austin S. Nuxoll. Characterization of Staphylococcus lugdunensis biofilms through ethyl methanesulfonate mutagenesis[J]. AIMS Microbiology, 2024, 10(4): 880-893. doi: 10.3934/microbiol.2024038
Staphylococcus lugdunensis is a coagulase-negative species responsible for a multitude of infections. These infections often resemble those caused by the more pathogenic staphylococcal species, Staphylococcus aureus, such as skin and soft tissue infections, prosthetic joint infections, and infective endocarditis. Despite a high mortality rate and infections that differ from other coagulase-negative species, little is known regarding S. lugdunensis pathogenesis. The objective of this study is to identify the essential factors for biofilm formation in S. lugdunensis. S. lugdunensis was mutagenized through ethyl methanesulfonate (EMS) exposure, and the individual cells were separated using a cell sorter and examined for biofilm formation at 8 hr and 24 hr timepoints. Mutations that resulted in either increased or decreased biofilm formation were sequenced to identify the genes responsible for the respective phenotypes. A mutation within the S. lugdunensis surface protein A (slsA) gene was common among all of the low biofilm formers, thus suggesting that high expression of this protein is important in biofilm formation. However, other mutations common among the mutants with decreased biofilm formation were in the putative divalent cation transport gene, mgtE. Conversely, a mutation in the gene that codes for the von Willebrand factor binding protein, vwbl, was common among the mutants with increased biofilm formation. Following proteinase K treatment, a significant dispersal of the S. lugdunensis biofilm matrix occurred, thus confirming the presence of primarily protein-mediated biofilms; this is in agreement with previous S. lugdunensis studies. Additionally, all low biofilm formers exhibited decreased protein levels (1.95–2.77 fold change) within the biofilm matrix, while no difference was observed with extracellular DNA (eDNA) or polysaccharides. This study presents a unique methodology to identify genes that affect biofilm formation and sheds light on S. lugdunensis pathogenesis.

| [1] |
Heilbronner S, Foster TJ (2021) Staphylococcus lugdunensis: A skin commensal with invasive pathogenic potential. Clin Microbiol Rev 34. https://doi.org/10.1128/cmr.00205-20 
|
| [2] | Parthasarathy S, Shah S, Raja Sager A, et al. (2020) Staphylococcus lugdunensis: Review of epidemiology, complications, and treatment. Cureus 12: e8801. https://doi.org/10.7759/cureus.8801 |
| [3] |
Liu PY, Huang YF, Tang CW, et al. (2010) Staphylococcus lugdunensis infective endocarditis: A literature review and analysis of risk factors. J Microbiol Immunol Infect 43: 478-484. https://doi.org/10.1016/S1684-1182(10)60074-6
|
| [4] |
Sandre RM, Shafran SD (1996) Infective endocarditis: Review of 135 cases over 9 years. Clin Infect Dis 22: 276-286. https://doi.org/10.1093/clinids/22.2.276
|
| [5] |
Karygianni L, Ren Z, Koo H, et al. (2020) Biofilm matrixome: Extracellular components in structured microbial communities. Trends Microbiol 28: 668-681. https://doi.org/10.1016/j.tim.2020.03.016
|
| [6] |
Theis TJ, Daubert TA, Kluthe KE, et al. (2023) Staphylococcus aureus persisters are associated with reduced clearance in a catheter-associated biofilm infection. Front Cell Infect Microbiol 13: 1178526. https://doi.org/10.3389/fcimb.2023.1178526
|
| [7] |
Otto M (2018) Staphylococcal biofilms. Microbiol Spectr 6. https://doi.org/10.1128/microbiolspec.gpp3-0023-2018
|
| [8] |
Sionov RV, Steinberg D (2022) Targeting the holy triangle of quorum sensing, biofilm formation, and antibiotic resistance in pathogenic bacteria. Microorganisms 10: 1239. https://doi.org/10.3390/microorganisms10061239
|
| [9] |
Hernandez-Cuellar E, Tsuchiya K, Valle-Rios R, et al. (2023) Differences in biofilm formation by methicillin-resistant and methicillin-susceptible Staphylococcus aureus strains. Diseases 11: 160. https://doi.org/10.3390/diseases11040160
|
| [10] |
Heilbronner S, Holden MT, van Tonder A, et al. (2011) Genome sequence of Staphylococcus lugdunensis N920143 allows identification of putative colonization and virulence factors. FEMS Microbiol Lett 322: 60-67. https://doi.org/10.1111/j.1574-6968.2011.02339.x
|
| [11] |
Herbert S, Ziebandt AK, Ohlsen K, et al. (2010) Repair of global regulators in Staphylococcus aureus 8325 and comparative analysis with other clinical isolates. Infect Immun 78: 2877-2889. https://doi.org/10.1128/iai.00088-10
|
| [12] |
Mack D, Siemssen N, Laufs R (1992) Parallel induction by glucose of adherence and a polysaccharide antigen specific for plastic-adherent Staphylococcus epidermidis: Evidence for functional relation to intercellular adhesion. Infect Immun 60: 2048-2057. https://doi.org/10.1128/iai.60.5.2048-2057.1992
|
| [13] |
McCarthy H, Rudkin JK, Black NS, et al. (2015) Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front Cell Infect Microbiol 5: 1. https://doi.org/10.3389/fcimb.2015.00001
|
| [14] |
Khodaparast L, Khodaparast L, Shahrooei M, et al. (2016) The possible role of Staphylococcus epidermidis LPxTG surface protein SesC in biofilm formation. PLoS One 11: e0146704. https://doi.org/10.1371/journal.pone.0146704
|
| [15] |
Assefa M, Amare A (2022) Biofilm-associated multi-drug resistance in hospital-acquired infections: A review. Infect Drug Resist 15: 5061-5068. https://doi.org/10.2147/IDR.S379502
|
| [16] |
Jamal M, Ahmad W, Andleeb S, et al. (2018) Bacterial biofilm and associated infections. J Chin Med Assoc 81: 7-11. https://doi.org/10.1016/j.jcma.2017.07.012
|
| [17] |
Lewis K (2010) Persister cells. Annu Rev Microbiol 64: 357-372. https://doi.org/10.1146/annurev.micro.112408.134306
|
| [18] |
Lourtet-Hascoet J, Bicart-See A, Felice MP, et al. (2016) Staphylococcus lugdunensis, a serious pathogen in periprosthetic joint infections: Comparison to Staphylococcus aureus and Staphylococcus epidermidis. Int J Infect Dis 51: 56-61. https://doi.org/10.1016/j.ijid.2016.08.007
|
| [19] |
Schroeder K, Jularic M, Horsburgh SM, et al. (2009) Molecular characterization of a novel Staphylococcus aureus surface protein (SasC) involved in cell aggregation and biofilm accumulation. PLoS One 4: e7567. https://doi.org/10.1371/journal.pone.0007567
|
| [20] | Corrigan RM, Rigby D, Handley P, et al. (2007) The role of Staphylococcus aureus surface protein SasG in adherence and biofilm formation. Microbiolog 153: 2435-2446. https://doi.org/10.1099/mic.0.2007/006676-0 |
| [21] |
Cucarella C, Solano C, Valle J, et al. (2001) Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J Bacteriol 183: 2888-2896. https://doi.org/10.1128/jb.183.9.2888-2896.2001
|
| [22] |
Rohde H, Burdelski C, Bartscht K, et al. (2005) Induction of Staphylococcus epidermidis biofilm formation via proteolytic processing of the accumulation-associated protein by staphylococcal and host proteases. Mol Microbiol 55: 1883-1895. https://doi.org/10.1111/j.1365-2958.2005.04515.x
|
| [23] |
Frank KL, Patel R (2007) Poly-N-acetylglucosamine is not a major component of the extracellular matrix in biofilms formed by icaADBC-positive Staphylococcus lugdunensis isolates. Infect Immun 75: 4728-4742. https://doi.org/10.1128/iai.00640-07
|
| [24] |
Missineo A, Di Poto A, Geoghegan JA, et al. (2014) IsdC from Staphylococcus lugdunensis induces biofilm formation under low-iron growth conditions. Infect Immun 82: 2448-2459. https://doi.org/10.1128/iai.01542-14
|
| [25] |
Steiger EL, Muelli JR, Braissant O, et al. (2020) Effect of divalent ions on cariogenic biofilm formation. BMC Microbiol 20: 1-11. https://doi.org/10.1186/s12866-020-01973-7
|
| [26] |
Yang J, Wei W, Pi S, et al. (2015) Competitive adsorption of heavy metals by extracellular polymeric substances extracted from Klebsiella sp. J1. Bioresour Technol 196: 533-539. https://doi.org/10.1016/j.biortech.2015.08.011
|
| [27] |
Sarkisova S, Patrauchan MA, Berglund D, et al. (2005) Calcium-induced virulence factors associated with the extracellular matrix of mucoid Pseudomonas aeruginosa biofilms. J Bacteriol 187: 4327-4337. https://doi.org/10.1128/jb.187.13.4327-4337.2005
|
| [28] |
Zhao X, Duan X, Dai Y, et al. (2020) Mycobacterium Von Willebrand factor protein MSMEG_3641 is involved in biofilm formation and intracellular survival. Future Microbiol 15: 1033-1044. https://doi.org/10.2217/fmb-2020-0064
|
| [29] |
Motta C, Pellegrini A, Camaione S, et al. (2023) von Willebrand factor-binding protein (vWbp)-activated factor XIII and transglutaminase 2 (TG2) promote cross-linking between FnBPA from Staphylococcus aureus and fibrinogen. Sci Rep 13: 11683. https://doi.org/10.1038/s41598-023-38972-3
|
| [30] |
Viljoen A, Viela F, Mathelie-Guinlet M, et al. (2021) Staphylococcus aureus vWF-binding protein triggers a strong interaction between clumping factor A and host vWF. Commun Biol 4: 453. https://doi.org/10.1038/s42003-021-01986-6
|
| [31] |
Viela F, Prystopiuk V, Leprince A, et al. (2019) Staphylococcus aureus protein A to von Willebrand factor is regulated by mechanical force. MBio 10. https://doi.org/10.1128/mbio.00555-19
|
| [32] |
Thomas S, Liu W, Arora S, et al. (2019) The complex fibrinogen interactions of the Staphylococcus aureus coagulases. Front Cell Infect Microbiol 9: 106. https://doi.org/10.3389/fcimb.2019.00106
|
| [33] |
Heilbronner S, Hanses F, Monk IR, et al. (2013) Sortase A promotes virulence in experimental Staphylococcus lugdunensis endocarditis. Microbiology 159: 2141-2152. https://doi.org/10.1099/mic.0.070292-0
|




Figures(5) / Tables(2)

McKenna J. Cruikshank, Justine M. Pitzer, Kimia Ameri, Caleb V. Rother, Kathryn Cooper, Austin S. Nuxoll. Characterization of Staphylococcus lugdunensis biofilms through ethyl methanesulfonate mutagenesis[J]. AIMS Microbiology, 2024, 10(4): 880-893. doi: 10.3934/microbiol.2024038







 DownLoad:
DownLoad: