Citation: Andrzej Maląg, Grzegorz Sobczak, Elżbieta Dąbrowska, Marian Teodorczyk. In-junction-plane beam divergence stabilization by lateral periodic structure in wide-stripe laser diodes[J]. AIMS Electronics and Electrical Engineering, 2019, 3(4): 370-381. doi: 10.3934/ElectrEng.2019.4.370

| [1] |
Larsson A, Salzman J, Mittelstein M, et al. (1986) Lateral coherence properties of broad-area semiconductor quantum well lasers. J Appl Phys 60: 66-68. doi: 10.1063/1.337629

|
| [2] |
Chow WW, Depatie D (1988) Filamentation in conventional double heterostructure and quantum well semiconductor lasers. IEEE J Quantum Elect 24: 1297-1301. doi: 10.1109/3.966
|
| [3] |
Chang-Haspain CJ, Kapon E, Colas E (1990) Spatial mode structure of index-guided broad-area quantum-well lasers. IEEE J Quantum Elect 26: 1713-1716. doi: 10.1109/3.60894
|
| [4] |
Dente GG (2001) Low confinement factors for suppressed filaments in semiconductors lasers. IEEE J Quantum Elect 37: 1650-1653. doi: 10.1109/3.970913
|
| [5] | Asatsuma T, Takiguchi Y, Frederico S, et al. (2006) Successive phase change and stability of near-field patterns for broad-area laser diodes. High-Power Diode Laser Technology and Applications IV 6104: 61040C. |
| [6] |
Tachikawa T, Takimoto S, Shogenji R, at al. (2010) Dynamics of broad-area semiconductor lasers with short optical feedback. IEEE J Quantum Elect 46: 140-149. doi: 10.1109/JQE.2009.2031122
|
| [7] | Wenzel H, Crump P, Ekhteraei H, et al. (2011) Theoretical and experimental analysis of the lateral modes of high-power broad-area lasers. 2011 Numerical Simulation of Optoelectronics Devices: 143-144. |
| [8] |
Crump P, Boldicke S, Schultz CM, et al. (2012) Experimental and theoretical analysis of the dominant lateral waveguiding mechanism in 975 nm high power broad area diode lasers. Semicond Sci Tech 27: 045001. doi: 10.1088/0268-1242/27/4/045001
|
| [9] |
Piprek J (2013) Self-consistent far-field blooming analysis for high-power Fabry-Perot laser diodes. Physics and Simulation of Optoelectronic Devices XXI 8619: 861910. doi: 10.1117/12.2004665
|
| [10] |
Winterfeldt M, Crump P, Wenzel H, et al. (2014) Experimental investigation of factors limiting slow axis beam quality in 9xx nm high power broad area diode lasers. J Appl Phys 116: 063103. doi: 10.1063/1.4892567
|
| [11] | Ripper JE, Paoli TL (1970) Optical coupling of adjacent stripe-geometry junction lasers. Appl Phys Lett 17: 371-373. |
| [12] | Scifres DR, Lindstrom C, Burnham RD, et al. (1983) Phase-locked (GaAl)As Laser diode emitting 2.6 W CW from a single mirror. Electron Lett 19: 169-171. |
| [13] |
Marshall W, Katz J (1986) Direct analysis of gain-guided phase-locked semiconductor laser arrays. IEEE J Quantum Elect 22: 827-832. doi: 10.1109/JQE.1986.1073064
|
| [14] |
Chaly VP, Karpov SY, Ter-Martirosyan AL, et al. (1996) Mechanisms of optical confinement in phase-locked laser arrays. Semicond Sci Tech 11: 372-379. doi: 10.1088/0268-1242/11/3/016
|
| [15] |
Maximov MV, Shernyakov YM, Novikov II, et al. (2008) High-power low-beam divergence edge-emitting semiconductor lasers with 1- and 20D photonic bandgap crystal waveguide. IEEE J Sel Top Quant 14: 1113-1122. doi: 10.1109/JSTQE.2008.916534
|
| [16] | Sobczak G, Maląg A (2012) Optimization of phase-locked arrays geometry for high-brightness laser systems. Opto-electron Rev 20: 134-137. |
| [17] |
Liu L, Zhang J, Wang Y, et al. (2012) 500-mW CW single-lobe emission from laterally coupled photonic crystal laser arrays. IEEE Photonic Tech Lett 24: 1667-1669. doi: 10.1109/LPT.2012.2209635
|
| [18] |
Borruel L, Sujecki S, Rodriguez D, et al. (2003) Beam filamentation and maximum optical power in high brightness tapered lasers. Physics and Simulation of Optoelectronic Devices XI 4986: 423-431. doi: 10.1117/12.474375
|
| [19] |
Wenzel H, Crump P, Fricke J, et al. (2013) Suppression of high-order lateral modes in broad-area diode lasers by resonant anti guiding. IEEE J Quantum Elect 49: 1102-1108. doi: 10.1109/JQE.2013.2288002
|
| [20] |
Leidner JP, Marciante JR (2012) Beam quality improvement in broad-area semiconductor lasers via evanescent spatial filtering. IEEE J Quantum Elect 48: 1269-1274. doi: 10.1109/JQE.2012.2207881
|
| [21] | Qiao Z, Zhang S, Gao X, et al. (2010) Broad area semiconductor lasers with tailored gain. Laser Physics and Laser Technologies (RCSLPLT) and 2010 Academic Symposium on Optoelectronics Technology (ASOT): 90-92. |
| [22] | Sun W, Pathak R, Campbell G, et al. (2013) Higher brightness laser diodes with smaller slow axis divergence. High-Power Diode Laser Technology and Applications XI 8605: 8605D. |
| [23] |
Piprek J (2013) Inverse thermal lens effects on the far-field blooming of broad area laser diodes. IEEE Photonic Tech Lett 25: 958-960. doi: 10.1109/LPT.2013.2255590
|
| [24] |
Takimoto S, Tachikawa T, Shogenji R, et al. (2009) Control of spatio-temporal dynamics of broad-area semiconductor lasers by strong optical injection. IEEE Photonnic Tech Lett 21: 1051-1053. doi: 10.1109/LPT.2009.2022181
|
| [25] |
An H, Xiong Y, Jiang CL, et al. (2014) Methods for slow axis beam quality improvement of high power broad area diode lasers. High-Power Diode Laser Technology and Applications XII 8965: 89650U. doi: 10.1117/12.2040986
|
| [26] | Yanson D, Levy M, Peleg O, et al. (2015) Low-NA fiber laser pumps powered by high-brightness single emitters. High-Power Diode Laser Technology and Applications XIII 9348: 934806. |
| [27] | Kanskar M, Bao L, Chen Z, et al. (2015) High brightness diodes & fiber-coupled modules. High-Power Diode Laser Technology and Applications XIII 9348: 934804. |
| [28] |
Winterfeldt M, Crump P, Knigge S, et al. (2015) High beam quality in broad area lasers via suppression of lateral carrier accumulation. IEEE Photonic Tech Lett 27: 1809-1812. doi: 10.1109/LPT.2015.2443186
|
| [29] | Liu G, Li J, Fan L, et al. (2017) High power single lateral mode 1050 nm laser diode bar. High-Power Diode Laser Technology XV 10086: 10086Y. |
| [30] |
Rieprich J, Winterfeldt M, Kerneke R, et al. (2018) Chip-carrier thermal barrier and its impact on lateral thermal lens profile and beam parameter product in high power broad area lasers. J Appl Phys 123: 125703. doi: 10.1063/1.5004503
|
| [31] | Kanskar M, Bai C, Bao L, et al. (2019) High brightness diodes and 600W and 60% efficient fiber-coupled packages enabled by reduced-mode (REM) diodes. High-Power Diode Laser Technology XVII 10900: 109000H. |
| [32] | Yang JT, Kim Y, Lee JB, et al. (2019) Dependence of high-power laser diode performance on emitter width. High-Power Diode Laser Technology XVII 10900: 109000N. |
| [33] |
Sobczak G, Dąbrowska E, Teodorczyk M, et al. (2014) Improvement of the lateral mode stability in high-power laser diodes by multi-stripe-gain distribution. IEEE J Quantum Elect 50: 890-897. doi: 10.1109/JQE.2014.2359237
|
| [34] |
Wang T, Tong C, Wang L, et al. (2016) Injection-insensitive lateral divergence in broad-area diode lasers achieved by spatial current modulation. Appl Phys Express 9: 112102. doi: 10.7567/APEX.9.112102
|
| [35] |
Maląg A, Dąbrowska E, Teodorczyk M, et al. (2012) Asymmetric heterostructure with reduced distance from active region to heatsink for 810-nm range high-power laser diodes. IEEE J Quantum Elect 48: 465-471. doi: 10.1109/JQE.2012.2184741
|
| [36] |
Tachikawa T, Takimoto S, Shogenji R, et al. (2010) Dynamics of broad-area semiconductor lasers with short optical feedback. IEEE J Quantum Elect 46: 140-148. doi: 10.1109/JQE.2009.2031122
|
| [37] | Sobczak G (2015) Improvement of the quality of the beam emitted by edge-emitting laser diodes through the lateral periodicity of heterostructure waveguide. Ph.D. Thesis. |
| [38] |
Kozłowska A, Maląg A, Dąbrowska E, et al. (2012) Thermal properties of high-power diode lasers investigated by means of high resolution thermography. Mat Sci Eng B 177: 1268-1272. doi: 10.1016/j.mseb.2012.02.024
|
| [39] |
Talghader J, Smith JS (1995) Thermal-dependence of the refractive-index of GaAs and AlAs measured using semiconductor multilayer optical cavities. Appl Phys Lett 66: 335-337. doi: 10.1063/1.114204
|
| [40] | Maląg A, Sobczak G, Dąbrowska E (2018) Emitted beam stabilization in junction plane by lateral periodic structure in laser diodes emitting at 980 nm. Laser Technology 2018: Progress and Applications of Lasers 10974: 1097404. |
| [41] | Martin D, Della Casa P, Adam T, et al. (2019) Current spreading suppression by O- and Si-implantation in high power broad area diode lasers. High-Power Diode Laser Technology XVII 10900: 109000M. |
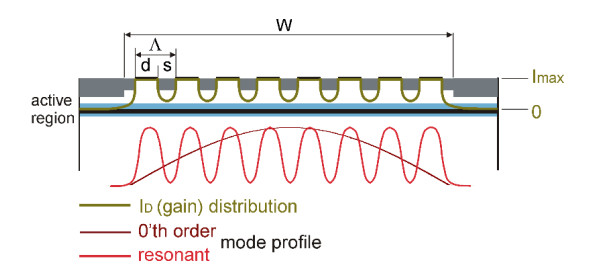
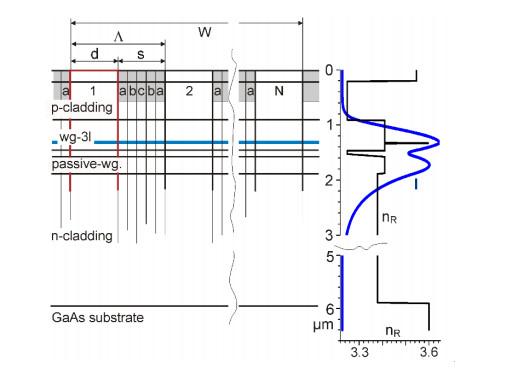
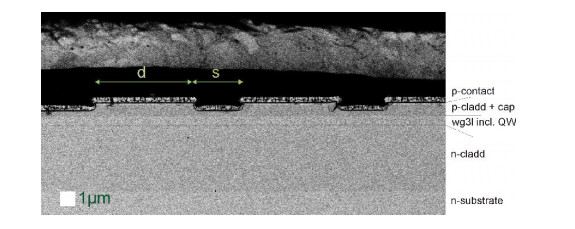
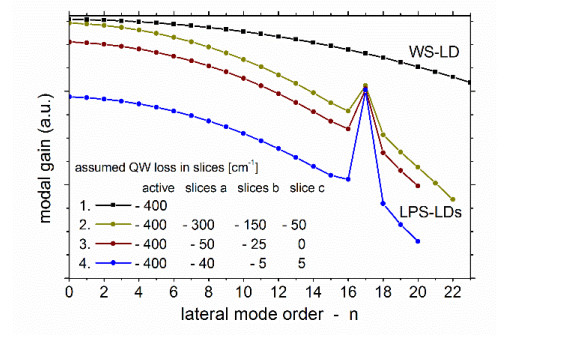
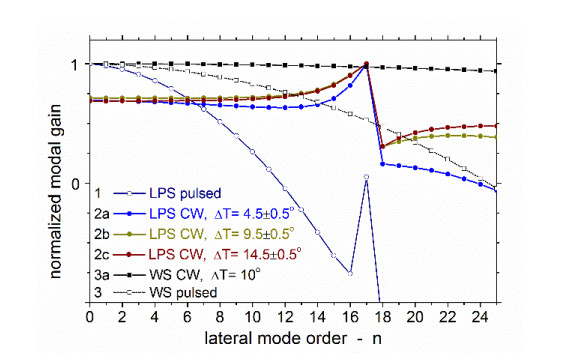
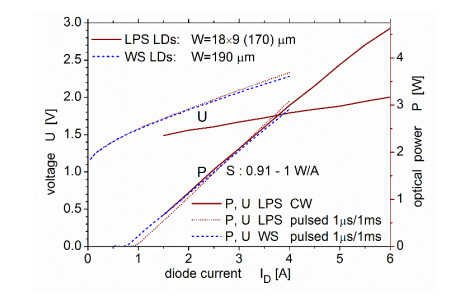
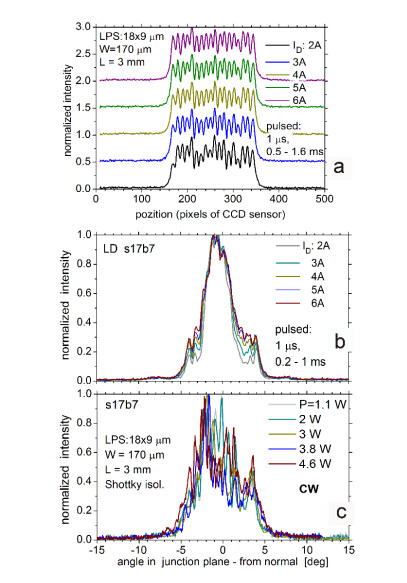
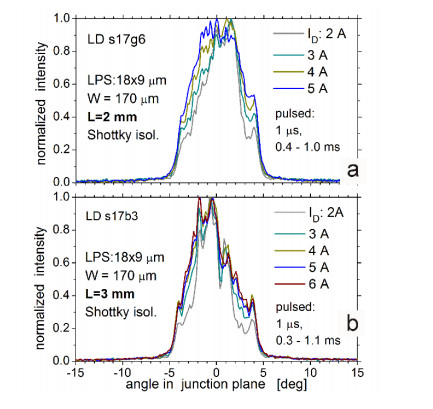
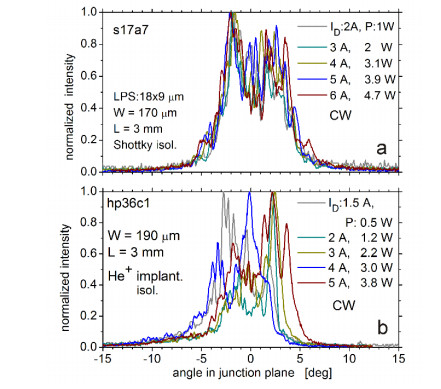
Figures(9)

Andrzej Maląg, Grzegorz Sobczak, Elżbieta Dąbrowska, Marian Teodorczyk. In-junction-plane beam divergence stabilization by lateral periodic structure in wide-stripe laser diodes[J]. AIMS Electronics and Electrical Engineering, 2019, 3(4): 370-381. doi: 10.3934/ElectrEng.2019.4.370







 DownLoad:
DownLoad: