Citation: Nguyen Hoai Nguyen. HY5, an integrator of light and temperature signals in the regulation of anthocyanins biosynthesis in Arabidopsis[J]. AIMS Molecular Science, 2020, 7(2): 70-81. doi: 10.3934/molsci.2020005

| [1] | Crozier A, Jaganath IB, Clifford MN (2006) Phenols, polyphenols and tannins: an overview. Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet Oxford, UK: Blackwell Publishing Ltd, 1-24. |
| [2] |
Harborne JB, Williams CA (2000) Advances in flavonoid research since 1992. Phytochemistry 55: 481-504. doi: 10.1016/S0031-9422(00)00235-1

|
| [3] |
Wang LS, Stoner GD (2008) Anthocyanins and their role in cancer prevention. Cancer Lett 269: 281-290. doi: 10.1016/j.canlet.2008.05.020
|
| [4] |
Romagnolo DF, Selmin OI (2012) Flavonoids and cancer prevention: a review of the evidence. J Nutr Gerontol Geriatr 31: 206-238. doi: 10.1080/21551197.2012.702534
|
| [5] |
Chen AY, Chen YC (2013) A review of the dietary flavonoid, kaempferol on human health and cancer chemoprevention. Food Chem 138: 2099-2107. doi: 10.1016/j.foodchem.2012.11.139
|
| [6] |
Peer WA, Murphy AS (2007) Flavonoids and auxin transport: modulators or regulators? Trends Plant Sci 12: 556-563. doi: 10.1016/j.tplants.2007.10.003
|
| [7] |
Cotelle N (2001) Role of flavonoids in oxidative stress. Curr Top Med Chem 1: 569-590. doi: 10.2174/1568026013394750
|
| [8] |
Howard LR, Pandjaitan N, Morelock T, et al. (2002) Antioxidant capacity and phenolic content of spinach as affected by genetics and growing season. J Agric Food Chem 50: 5891-5896. doi: 10.1021/jf020507o
|
| [9] | Oh JE, Kim YH, Kim JH, et al. (2011) Enhanced level of anthocyanin leads to increased salt tolerance in Arabidopsis PAP1-D plants upon sucrose treatment. J Korean Soc Appl Bi 54: 79-88. |
| [10] |
Di Ferdinando M, Brunetti C, Fini A, et al. (2012) Flavonoids as antioxidants in plants under abiotic stresses. Abiotic stress responses in plants: metabolism productivity and sustainability New York: Springer, 159-179. doi: 10.1007/978-1-4614-0634-1_9
|
| [11] |
Nakabayashi R, Yonekura-Sakakibara K, Urano K, et al. (2014) Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J 77: 367-379. doi: 10.1111/tpj.12388
|
| [12] |
Nguyen NH, Kim JH, Kwon J, et al. (2016) Characterization of Arabidopsis thaliana FLAVONOL SYNTHASE 1 (FLS1)-overexpression plants in response to abiotic stress. Plant Physiol Biochem 103: 133-142. doi: 10.1016/j.plaphy.2016.03.010
|
| [13] |
Kuhn BM, Geisler M, Bigler L, et al. (2011) Flavonols accumulate asymmetrically and affect auxin transport in Arabidopsis. Plant Physiol 156: 585-595. doi: 10.1104/pp.111.175976
|
| [14] |
Petrussa E, Braidot E, Zancani M, et al. (2013) Plant flavonoids-biosynthesis, transport and involvement in stress responses. Int J Mol S 14: 14950-14973. doi: 10.3390/ijms140714950
|
| [15] |
Saslowsky DE, Ware U, Winkel BSJ (2005) Nuclear localization of flavonoid enzymes in Arabidopsis. J Bio Chem 280: 23735-23740. doi: 10.1074/jbc.M413506200
|
| [16] |
Tian L, Wan SB, Pan QH, et al. (2008) A novel plastid localization of chalcone synthase in developing grape berry. Plant Sci 175: 431-436. doi: 10.1016/j.plantsci.2008.03.012
|
| [17] |
Wang HL, Wang W, Zhang P, et al. (2010) Gene transcript accumulation, tissue and subcellular localization of anthocyanidin synthase (ANS) in developing grape berries. Plant Sci 179: 103-113. doi: 10.1016/j.plantsci.2010.04.002
|
| [18] |
Toda K, Kuroiwa H, Senthil K, et al. (2012) The soybean F3′H protein is localized to the tonoplast in the seed coat hilum. Planta 236: 79-89. doi: 10.1007/s00425-012-1590-5
|
| [19] |
Winkel-Shirley B (2002) Biosynthesis of flavonoids and effects of stress. Curr Opin Plant Biol 5: 218-223. doi: 10.1016/S1369-5266(02)00256-X
|
| [20] |
Fini A, Brunetti C, Di Ferdinando M, et al. (2011) Stress-induced flavonoid biosynthesis and the antioxidant machinery of plants. Plant Signal Behav 6: 709-711. doi: 10.4161/psb.6.5.15069
|
| [21] |
Nguyen NH, Jeong CY, Kang GH, et al. (2015) MYBD employed by HY5 increases anthocyanin accumulation via repression of MYBL2 in Arabidopsis. Plant J 84: 1192-1205. doi: 10.1111/tpj.13077
|
| [22] |
Wang Y, Wang Y, Song Z, et al. (2016) Repression of MYBL2 by both microRNA858a and HY5 leads to the activation of anthocyanin biosynthetic pathway in Arabidopsis. Mol Plant 9: 1395-1405. doi: 10.1016/j.molp.2016.07.003
|
| [23] |
Kim S, Hwang G, Lee S, et al. (2017) High ambient temperature represses anthocyanin biosynthesis through degradation of HY5. Front Plant Sci 8: 1787. doi: 10.3389/fpls.2017.01787
|
| [24] |
Vogt T (2010) Phenylpropanoid biosynthesis. Mol Plant 3: 2-20. doi: 10.1093/mp/ssp106
|
| [25] |
Holton TA, Cornish EC (1995) Genetics and Biochemistry of Anthocyanin Biosynthesis. Plant Cell 7: 1071-1083. doi: 10.2307/3870058
|
| [26] |
Tanaka Y, Sasaki N, Ohmiya A (2008) Biosynthesis of plant pigments: anthocyanins, betalains and carotenoids. Plant J 54: 733-749. doi: 10.1111/j.1365-313X.2008.03447.x
|
| [27] |
Rosinski JA, Atchley WR (1998) Molecular evolution of the Myb family of transcription factors: Evidence for polyphyletic origin. J Mol Evol 46: 74-83. doi: 10.1007/PL00006285
|
| [28] |
Jin HL, Martin C (1999) Multi-functionality and diversity within the plant MYB-gene family. Plant Mol Biol 41: 577-585. doi: 10.1023/A:1006319732410
|
| [29] |
Dubos C, Stracke R, Grotewold E, et al. (2010) MYB transcription factors in Arabidopsis. Trends Plant Sci 15: 573-581. doi: 10.1016/j.tplants.2010.06.005
|
| [30] |
Ogata K, Morikawa S, Nakamura H, et al. (1994) Solution structure of a specific DNA complex of the Myb DNA-binding domain with cooperative recognition helices. Cell 79: 639-648. doi: 10.1016/0092-8674(94)90549-5
|
| [31] |
Dias AP, Braun EL, McMullen MD, et al. (2003) Recently duplicated maize R2R3 Myb genes provide evidence for distinct mechanisms of evolutionary divergence after duplication. Plant Physiol 131: 610-620. doi: 10.1104/pp.012047
|
| [32] |
Matus JT, Aquea F, Arce-Johnson P (2008) Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across Vitis and Arabidopsis genomes. BMC Plant Biol 8: 83. doi: 10.1186/1471-2229-8-83
|
| [33] |
Nguyen NH, Cheong JJ (2018) AtMYB44 interacts with TOPLESS-RELATED corepressors to suppress Protein Phosphatase 2C gene transcription. Biochem Biophys Res Commun 499: 437-442. doi: 10.1016/j.bbrc.2018.11.057
|
| [34] |
Stracke R, Ishihara H, Barsch GHA, et al. (2007) Differential regulation of closely related R2R3-MYB transcription factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J 50: 660-677. doi: 10.1111/j.1365-313X.2007.03078.x
|
| [35] | Koornneef M (1990) Mutations affecting the testa color in Arabidopsis. Arabid Inf Serv 27: 1-4. |
| [36] | Meyerowitz EM, Bowman JL, Chang C, et al. (1990) RFLP map of Arabidopsis thaliana. Cold Spring Harbor New York: Cold Spring Harbor Laboratory Press, 98-99. |
| [37] | Shirley BW, Hanley S, Goodman HM (1992) Effects of ionizing radiation on a plant genome: analysis of two Arabidopsis transparent testa mutations. Plant Cell 4: 333-347. |
| [38] |
Focks N, Sagasser M, Weisshaar B, et al. (1999) Characterization of tt15, a novel transparent testa mutant of Arabidopsis thaliana (L.) Heynh. Planta 208: 352-357. doi: 10.1007/s004250050569
|
| [39] | Fuglevand G, Jackson JA, Jenkins GI (1996) UV-B, UV-A, and blue light signal transduction pathways interact synergistically to regulate chalcone synthase gene expression in Arabidopsis. Plant Cell 8: 2347-2357. |
| [40] |
Wade HK, Bibikova TN, Valentine WJ, et al. (2001) Interaction within a network of phytochrome, cryptochrome and UV-B phototransduction pathways regulate chalcone synthase gene expression in Arabidopsis leaf tissue. Plant J 25: 675-685. doi: 10.1046/j.1365-313x.2001.01001.x
|
| [41] |
Mehrtens F, Kranz H, Bednarek P, et al. (2005) The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol 138: 1083-1096. doi: 10.1104/pp.104.058032
|
| [42] |
Borevitz JO, Xia YJ, Blount J, et al. (2000) Activation tagging identifies a conserved MYB regulator of phenylpropanoid biosynthesis. Plant Cell 12: 2383-2393. doi: 10.1105/tpc.12.12.2383
|
| [43] |
Nguyen H, Kim JH, Hyun WY, et al. (2013) TTG1-mediated flavonols biosynthesis alleviates root growth inhibition in response to ABA. Plant Cell Rep 32: 503-514. doi: 10.1007/s00299-012-1382-1
|
| [44] |
Li C, Zhang B, Chen B, et al. (2018b) Site-specific phosphorylation of TRANSPARENT TESTA GLABRA1 mediates carbon partitioning in Arabidopsis seeds. Nat Commun 9: 571. doi: 10.1038/s41467-018-03013-5
|
| [45] |
Baudry A, Heim MA, Dubreucq B, et al. (2004) TT2, TT8, and TTG1 synergistically specify the expression of BANYULS and proanthocyanidin biosynthesis in Arabidopsis thaliana. Plant J 39: 366-380. doi: 10.1111/j.1365-313X.2004.02138.x
|
| [46] |
Xu W, Dubos C, Lepiniec L (2015) Transcriptional control of flavonoid biosynthesis by MYB-bHLH-WDR complexes. Trends Plant Sci 20: 176-185. doi: 10.1016/j.tplants.2014.12.001
|
| [47] |
Dubos C, Le Gourrierec J, Baudry A, et al. (2008) MYBL2 is a new regulator of flavonoid biosynthesis in Arabidopsis thaliana. Plant J 55: 940-953. doi: 10.1111/j.1365-313X.2008.03564.x
|
| [48] |
Matsui K, Umemura Y, Ohme-Takagi M (2008) AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. Plant J 55: 954-967. doi: 10.1111/j.1365-313X.2008.03565.x
|
| [49] |
Gou JY, Felippes FF, Liu CJ, et al. (2011) Negative regulation of anthocyanin biosynthesis in arabidopsis by a miR156-Targeted SPL transcription factor. Plant Cell 23: 1512-1522. doi: 10.1105/tpc.111.084525
|
| [50] |
Shin J, Park E, Choi G (2007) PIF3 regulates anthocyanin biosynthesis in an HY5-dependent in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis. Plant J 50: 933. doi: 10.1111/j.1365-313X.2007.03178.x
|
| [51] |
Vandenbussche F, Habricot Y, Condiff AS, et al. (2007) HY5 is a point of convergence between cryptochrome and cytokinin signalling pathways in Arabidopsis thaliana. Plant J 49: 428-441. doi: 10.1111/j.1365-313X.2006.02973.x
|
| [52] |
Shin DH, Choi M, Kim K, et al. (2013) HY5 regulates anthocyanin biosynthesis by inducing the transcriptional activation of the MYB75/PAP1 transcription factor in Arabidopsis. FEBS Lett 587: 1543-1547. doi: 10.1016/j.febslet.2013.03.037
|
| [53] |
Pietta PG (2000) Flavonoids as antioxidants. J Nat Prod 63: 1035-1042. doi: 10.1021/np9904509
|
| [54] |
Potapovich AI, Kostyuk VA (2003) Comparative study of antioxidant properties and cytoprotective activity of flavonoids. Biochemistry (Mosc) 68: 514-519. doi: 10.1023/A:1023947424341
|
| [55] |
Maier A, Schrader A, Kokkelink L, et al. (2013) Light and the E3 ubiquitin ligase COP1/SPA control the protein stability of the MYB transcription factors PAP1 and PAP2 involved in anthocyanin accumulation in Arabidopsis. Plant J 74: 638-651. doi: 10.1111/tpj.12153
|
| [56] |
Das PK, Geul B, Choi SB, et al. (2011) Photosynthesis-dependent anthocyanin pigmentation in Arabidopsis. Plant Signal Behav 6: 23-25. doi: 10.4161/psb.6.1.14082
|
| [57] |
Zoratti L, Karppinen K, Escobar AL, et al. (2014) Light-controlled flavonoid biosynthesis in fruits. Front Plant Sci 5: 534. doi: 10.3389/fpls.2014.00534
|
| [58] |
Saure MC (1990) External control of anthocyanin formation in apple. Sci Hortic 42: 181-218. doi: 10.1016/0304-4238(90)90082-P
|
| [59] |
Chen DQ, Li ZY, Pan RC, et al. (2006) Anthocyanin accumulation mediated by blue light and cytokinin in Arabidopsis seedlings. J Integr Plant Biol 48: 420-425. doi: 10.1111/j.1744-7909.2006.00234.x
|
| [60] |
Cao SF, Hu ZC, Zheng YH, et al. (2010) Effect of BTH on anthocyanin content and activities of related enzymes in strawberry after harvest. J Agric Food Chem 58: 5801-5805. doi: 10.1021/jf100742v
|
| [61] |
Kadomura-Ishikawa Y, Miyawaki K, Noji S, et al. (2013) Phototropin 2 is involved in blue light-induced anthocyanin accumulation in Fragaria x ananassa fruits. J Plant Res 126: 847-857. doi: 10.1007/s10265-013-0582-2
|
| [62] |
Xu F, Cao SF, Shi LY, et al. (2014) Blue light irradiation affects anthocyanin content and enzyme activities involved in postharvest strawberry fruit. J Agric Food Chem 62: 4778-4783. doi: 10.1021/jf501120u
|
| [63] |
Catala R, Medina J, Salinas J (2011) Integration of low temperature and light signaling during cold acclimation response in Arabidopsis. Proc Natl Acad Sci USA 108: 16475-16480. doi: 10.1073/pnas.1107161108
|
| [64] |
Mori K, Sugaya S, Gemma H (2005) Decreased anthocyanin biosynthesis in grape berries grown under elevated night temperature condition. Sci Hortic-Amsterdam 105: 319-330. doi: 10.1016/j.scienta.2005.01.032
|
| [65] | Yamane T, Jeong ST, Goto-Yamamoto N, et al. (2006) Effects of temperature on anthocyanin biosynthesis in grape berry skins. Am J Enol Viticult 57: 54-59. |
| [66] |
Koornneef M, Rolff E, Spruit CJP (1980) Genetic control of light-inhibited hypocotyl elongation in Arabidopsis thaliana (L) Heynh. Zeitschrift für Pflanzenphysiologie 100: 147-160. doi: 10.1016/S0044-328X(80)80208-X
|
| [67] |
Gangappa SN, Botto JF (2016) The multifaceted roles of HY5 in plant growth and development. Mol Plant 9: 1353-1365. doi: 10.1016/j.molp.2016.07.002
|
| [68] |
Oyama T, Shimura Y, Okada K (1997) The Arabidopsis HY5 gene encodes a bZIP protein that regulates stimulus-induced development of root and hypocotyl. Genes Dev 11: 2983-2995. doi: 10.1101/gad.11.22.2983
|
| [69] |
Ang LH, Chattopadhyay S, Wei N, et al. (1998) Molecular interaction between COP1 and HY5 defines a regulatory switch for light control of Arabidopsis development. Mol Cell 1: 213-222. doi: 10.1016/S1097-2765(00)80022-2
|
| [70] |
Laubinger S, Fittinghoff K, Hoecker U (2004) The SPA quartet: a family of WD-repeat proteins with a central role in suppression of photomorphogenesis in Arabidopsis. Plant Cell 16: 2293-2306. doi: 10.1105/tpc.104.024216
|
| [71] |
Lau OS, Deng XW (2012) The photomorphogenic repressors COP1 and DET1: 20 years later. Trends Plant Sci 17: 584-593. doi: 10.1016/j.tplants.2012.05.004
|
| [72] |
Podolec R, Ulm R (2018) Photoreceptor-mediated regulation of the COP1/SPA E3 ubiquitin ligase. Curr Opin Plant Biol 45: 18-25. doi: 10.1016/j.pbi.2018.04.018
|
| [73] |
Chattopadhyay S, Ang LH, Puente P, et al. (1998) Arabidopsis bZIP protein HY5 directly interacts with light-responsive promoters in mediating light control of gene expression. Plant Cell 10: 673-683. doi: 10.1105/tpc.10.5.673
|
| [74] |
Osterlund MT, Hardtke CS, Wei N, et al. (2000) Targeted destabilization of HY5 during light-regulated development of Arabidopsis. Nature 405: 462-466. doi: 10.1038/35013076
|
| [75] |
Li B, Gao K, Ren H, et al. (2018) Molecular mechanisms governing plant responses to high temperatures. J Integr Plant Biol 60: 757-779. doi: 10.1111/jipb.12701
|
| [76] |
Park YJ, Lee HJ, Ha JH, et al. (2017) COP1 conveys warm temperature information to hypocotyl thermomorphogenesis. New Phytol 215: 269-280. doi: 10.1111/nph.14581
|
| [77] | Stracke R, Favory JJ, Gruber H, et al. (2010) The Arabidopsis bZIP transcription factor HY5 regulates expression of the PFG1/MYB12 gene in response to light and ultraviolet-B radiation. Plant Cell Environ 33: 88-103. |
| [78] |
Zhang H, He H, Wang X, et al. (2011) Genome-wide mapping of the HY5-mediated gene networks in Arabidopsis that involve both transcriptional and post-translational regulation. Plant J 65: 346-358. doi: 10.1111/j.1365-313X.2010.04426.x
|
| [79] |
Burko Y, Seluzicki A, Zander M, et al. (2020) Chimeric activators and repressors define HY5 activity and reveal a light-regulated feedback mechanism. Plant Cell 32: 967-983. doi: 10.1105/tpc.19.00772
|
| [80] |
Ram H, Priya P, Jain M, et al. (2014) Genome-wide DNA binding of GBF1 is modulated by its heterodimerizing protein partners, HY5 and HYH. Mol Plant 7: 448-451. doi: 10.1093/mp/sst143
|
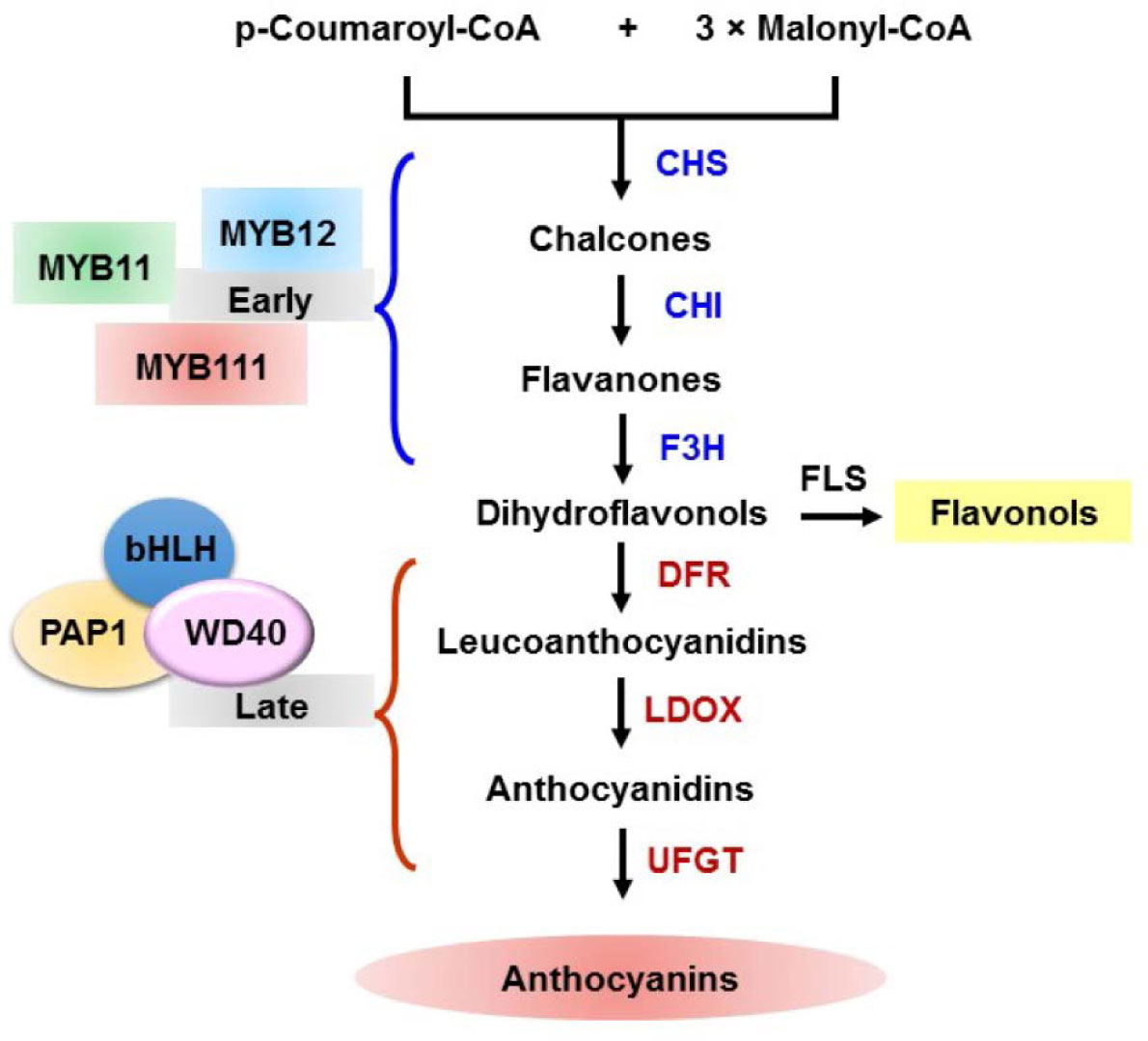
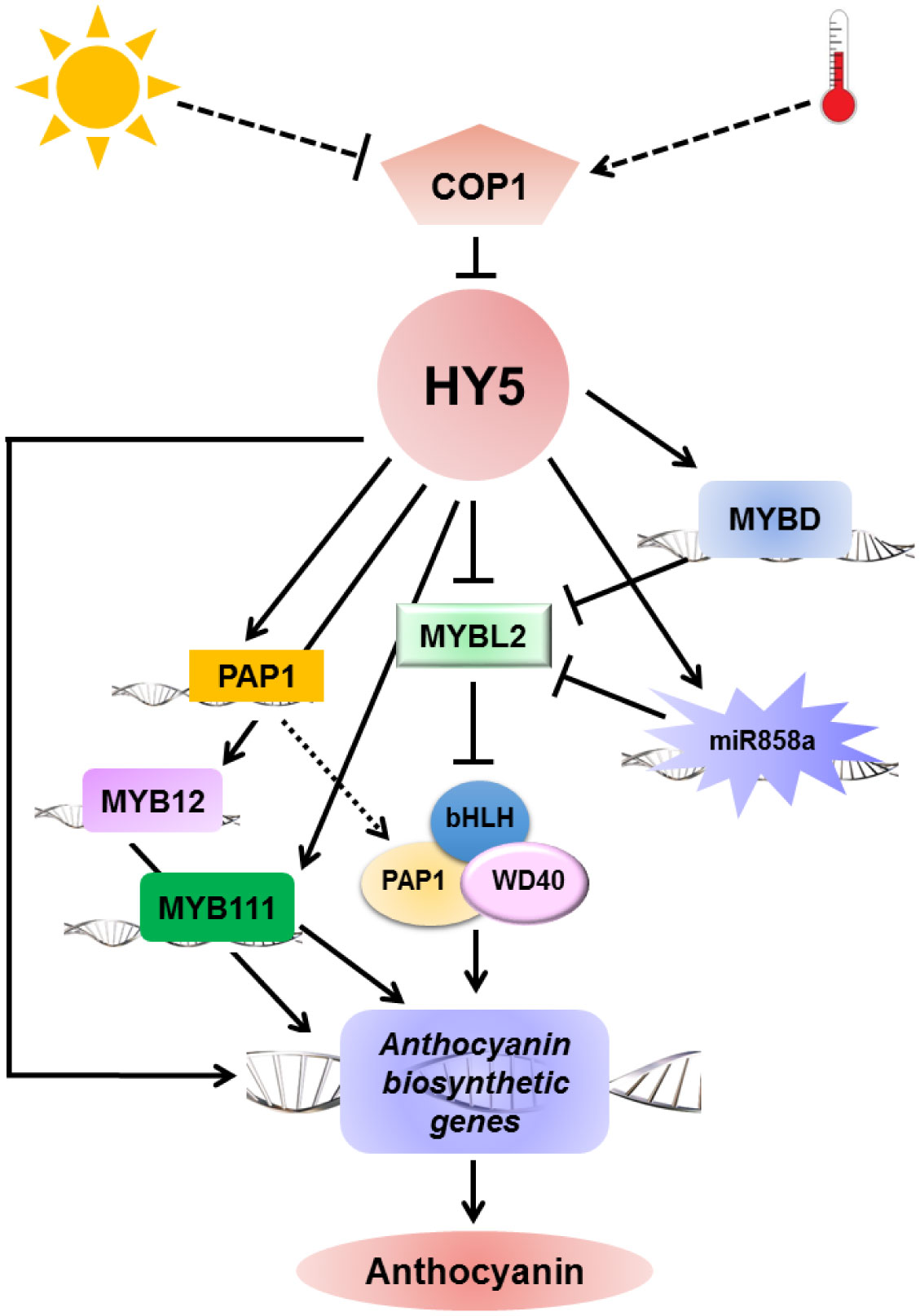
Figures(2)

Nguyen Hoai Nguyen. HY5, an integrator of light and temperature signals in the regulation of anthocyanins biosynthesis in Arabidopsis[J]. AIMS Molecular Science, 2020, 7(2): 70-81. doi: 10.3934/molsci.2020005







 DownLoad:
DownLoad: