Citation: Marcelo F. Pompelli, José N. B. Santos, Marcos A. Santos. Estimating leaf area of Jatropha nana through non-destructive allometric models[J]. AIMS Environmental Science, 2019, 6(2): 59-76. doi: 10.3934/environsci.2019.2.59

| [1] | Sabandar CW, Ahmat N, Jaafar FM, et al. (2013) Medicinal property, phytochemistry and pharmacology of several Jatropha species (Euphorbiaceae): a review. Phytochemistry, 85, 7–29. |
| [2] | Balakrishnan NP, Chakrabarty T. (2017) The family Euphorbiaceae in India: a synopsis of its profile, taxonomy and bibliography, Bishen Singh Mahendra Pal Singh, Dehradun. |
| [3] | Bahadur B, Sujatha M, Carels N Jatropha.(2012) Challenges for a New Energy Crop: Volume 2: Genetic Improvement and Biotechnology, Springer Science & Business Media, New York. |
| [4] | Nerlekar AN. (2015) Lectotypification of Jatropha nana (Euphorbiaceae) with notes on its threats, and the status of Jatropha nana var. benghalensis. Phytotaxa, 213, 155–158. |
| [5] | Banerji R. (1991) Jatropha nana seed oil for energy. Biomass Bioenerg, 1, 247–247. |
| [6] | Kirtikar KR, Basu BD. (1918) Indian Medicinal Plants Vol-3., Bishen Singh Mahendra Pal Singh, Dehradun. |
| [7] | Mishra DK, Singh NP. (2001) Endemic and Threatened plants of Maharashtra, Botanical Survey of India, Calcutta. |
| [8] | Nerlekar AN, Kumar A, Venu P. (2016) Jatropha nana. The IUCN Red List of Threatened Species 2016. In, Vol 2017. |
| [9] | Asner GP, Scurlock JMO, Hicke JA.(2003) Global synthesis of leaf area index observations: implications for ecological and remote sensing studies. Global Ecol Biogeogr, 12, 191–205. |
| [10] | Blanco FF, Folegatti MV. (2005) Estimation of leaf area for greenhouse cucumber by linear measurements under salinity and grafting. Sci Agr, 62, 305–309. |
| [11] | Cristofori V, Rouphael Y, Mendoza-de Gyves E, et al. (2007) A simple model for estimating leaf area of hazelnut from linear measurements. Sci Hort, 113, 221–225. |
| [12] | Bréda NJJ. (2003) Ground-based measurements of leaf area index: a review of methods, instruments and current controversies. J Exp Bot, 54, 2403–2417. |
| [13] | Díaz S, Kattge J, Cornelissen JHC, et al. (2016) The global spectrum of plant form and function. Nature, 529, 167–171. |
| [14] | Koyama K, Kikuzawa K. (2009) Is whole‐plant photosynthetic rate proportional to leaf area? A test of scalings and a logistic equation by leaf demography census. Am Nat, 173, 640–649. |
| [15] | Pompelli MF, Antunes WC, Ferreira DTRG, et al. (2012) Allometric models for non-destructive leaf area estimation of the Jatropha curcas. Biomass Bioenerg, 36, 77–85. |
| [16] | Peksen E. (2007) Non-destructive leaf area estimation model for faba bean (Vicia faba L.). Sci Hortic-Amsterdam, 2007, 113, 322–328. |
| [17] | Thomson VP, Cunningham SA, Ball MC, et al. (2003) Compensation for herbivory by Cucumis sativus through increased photosynthetic capacity and efficiency. Oecologia, 134, 167–175. |
| [18] | De Swart EAM, Groenwold R, Kanne HJ, Stam P, et al. (2004) Non-destructive estimation of leaf area for different plant ages and accessions of Capsicum annuum L. J Hortic Sci Biotech, 79, 764–770. |
| [19] | Antunes WC, Pompelli MF, Carretero DM, et al. (2008) Allometric models for non-destructive leaf area estimation in coffee (Coffea arabica and Coffea canephora). Ann Appl Biol, 153, 33–40. |
| [20] | Severino LS, Vale LS, Beltrão NEM. (2007) A simple method for measurement of Jatropha curcas leaf area. Rev Bras Ol Fibros, 11, 9–14. |
| [21] | Achten WMJ, Maes WH, Reubens B, et al. (2010) Biomass production and allocation in Jatropha curcas L. seedlings under different levels of drought stress. Biomass Bioenerg, 34, 667–676. |
| [22] | Champion HG, Seth SK. (1968) A revised survey of the forest types of India, Government of India Pblication, Delhi. |
| [23] | Zuur AF, Elena NI, Elphick CS. (2010) A protocol for data exploration to avoid common statistical problems. Method Ecol Evol, 1, 3–14. |
| [24] | Durbin J, Watson GS. (1950) Testing for serial correlation in least squares regression I. Biometrika, 37, 409–428. |
| [25] | Akaike H. (1974) A new look at the statistical model identification. Transac Autom Control, 19, 716–723. |
| [26] | Walther BA, Moore JL. (2005) The concepts of bias, precision and accuracy, and their use in testing the performance of species richness estimators, with a literature review of estimator performance. Ecography, 28, 815–829. |
| [27] | Graybill FA. (2000) Theory and application of the linear model, Duxbury Press, Pacific Grove. |
| [28] | Ahmed N, Khan D. (2011) Leaf area estimation in Jatropha curcas L. Int J Biol Biotech, 8, 401–407. |
| [29] | Chatterjee S, Hadi AS. (2006) Regression Analysis by Example, John Wiley & Sons, Hoboken. |
| [30] | Beerling DJ, Fry JC. (1990) A comparison of the accuracy, variability and speed of five different methods for estimating leaf area. Ann Bot, 65, 483–488. |
| [31] | Campostrini E, Yamanishi OK. (2001) Estimation of papaya leaf area using the central vein length. Sci Agric, 58, 39–42. |
| [32] | Kandiannan K, Parthasarathy U, Krishnamurthy KS, et al. (2009) Modeling individual leaf area of ginger (Zingiber officinale Roscoe) using leaf length and width. Sci Hortic-Amsterdam, 120, 532–537. |
| [33] | Negash M, Starr M, Kanninen M, et al. (2013) Allometric equations for estimating aboveground biomass of Coffea arabica L. grown in the Rift Valley escarpment of Ethiopia. Agrofor Syst, 87, 953–966. |
| [34] | Kandiannan K, Kailassam C, Chandaragiri KK, et al. (2002) Allometric model for leaf area estimation in black pepper (Piper nigrum L.). J Agron Crop Sci, 188, 138–140. |
| [35] | Williams IIIL, Martinson TE. (2003) Nondestructive leaf area estimation of 'Niagara' and 'DeChaunac' grapevines. Sci Hortic-Amsterdam, 98, 493–498. |
| [36] | Bosco LC, Bergamaschi H, Cardoso LS, et al. (2012) Selection of regression models for estimating leaf area of 'Royal gala' and 'Fuji suprema' apple trees under hail net and in open sky. Rev Bras Frutic, 34, 504–514. |
| [37] | Sangoi L, Schmitt A, Zanin CG. (2007 )Área foliar e rendimento de grãos de híbridos de milho em diferentes populações de planta. Rev Bras Milho e Sorgo, 6, 263–271. |
| [38] | Souza LF, Santos JGD, Alexandrino E, et al. (2015) Practical and efficient method to estimate leaf area index in tropical forage grasses. Arch Zootec, 64, 83–85. |
| [39] | Peters CA. (2001) Statistics for Analysis of Experimental Data. In, Vol 2017. S.E. Powers, Ed., Champaign |
| [40] | Francis CA, Rutger JN, Palmer AFE. (1969) A rapid method for plant leaf area estimation in maize (Zea mays L.). Crop Sci, 9, 537–539. |
| [41] | Elsner EA, Jubb Jr, GL. (1988) Leaf area estimation of concord grape leaves from simple linear measurements. Am J Enol Vitic, 39, 95–97. |
| [42] | Lu HY, Lu CT, Wei ML, et al. (2004) Comparison of different models for nondestructive leaf area estimation in taro. Agron J, 96, 448–453. |
| [43] | Rouphael Y, Colla G, Fanasca S, et al. (2007) Leaf area estimation of sunflower leaves from simple linear measurements. Photosynthetica, 45, 306–308. |
| [44] | Kumar R. (2009) Calibration and validation of regression model for non-destructive leaf area estimation of saffron (Crocus sativus L.). Sci Hortic-Amsterdam, 122, 142–145. |
| [45] | Barros RS, Maestri M, Vieira M, et al. (1973) Determinação da área de folhas do café (Coffea arabica L. cv. 'Bourbon Amarelo'). Rev Ceres, 20, 44–52. |
| [46] | Kurt O, Uysal H, Uzun S. (2005) Non-destructive leaf area estimation of flax (Linun usitatissimum L.). Pak J Bot, 37, 837–841. |
| [47] | Serdar Ü, Demirsoy H. (2006) Non-destructive leaf area estimation in chestnut. Sci Hortic-Amsterdam, 108, 227–230. |
| [48] | Rouphael Y, Mouneimne AH. A I, et al. (2010) Modeling individual leaf area of rose (Rosa hybrida L.) based on leaf length and width measurement. Photosynthetica, 48, 9–15. |
| [49] | Keramatlou I, Sharifani M, Sabouri H, et al. (2015) A simple linear model for leaf area estimation in Persian walnut (Juglans regia L.). Sci Hort, 184, 36–39. |
| [50] | Souza MC, Amaral CL. (2015) Non-destructive linear model for leaf area estimation in Vernonia ferruginea Less. Braz J Biol, 75, 152–156. |
| [51] | Boisbunon A, Canu S, Fourdrinier D, et al. (2014) Akaike's information criterion, Cp and estimators of loss for elliptically symmetric distributions. Int Stat Rev, 82, 422–439. |
| [52] | Vasconcellos ES, Cruz CD, Regazzi AJ, et al. (2010) Regression models grouping for genotype adaptability and stability analysis. Pesq Agrop Bras, 45, 1357–1362. |
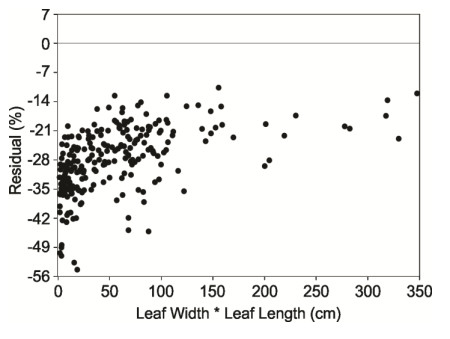
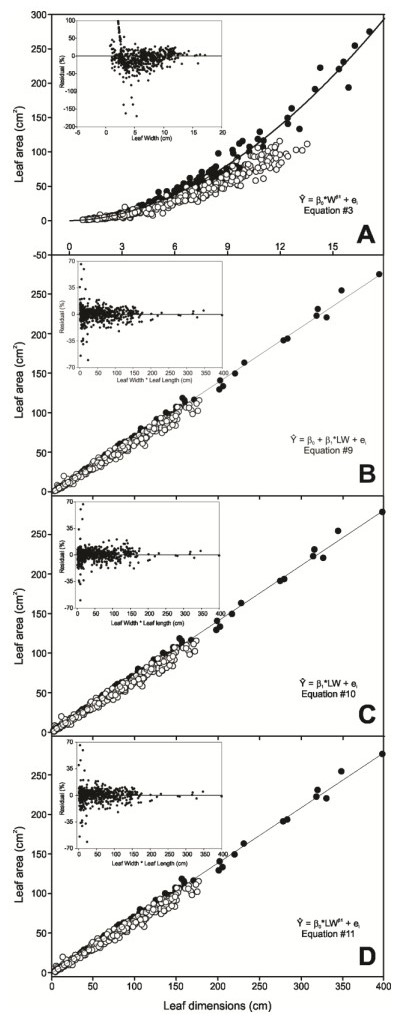
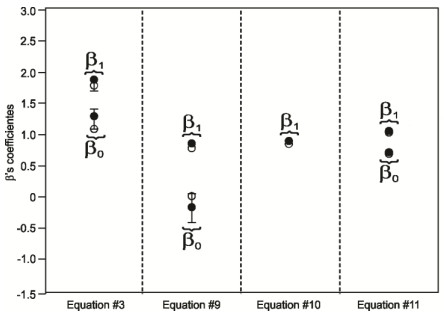
Figures(7) / Tables(4)

Marcelo F. Pompelli, José N. B. Santos, Marcos A. Santos. Estimating leaf area of Jatropha nana through non-destructive allometric models[J]. AIMS Environmental Science, 2019, 6(2): 59-76. doi: 10.3934/environsci.2019.2.59







 DownLoad:
DownLoad: