Citation: Robert Friedman. Themes of advanced information processing in the primate brain[J]. AIMS Neuroscience, 2020, 7(4): 373-388. doi: 10.3934/Neuroscience.2020023

| [1] | Ramon Y, Cajal S (1899) Textura del Sistema Nervioso del Hombre y de los Vertebrados trans Madrid: Nicolas Moya. |
| [2] | Brodmann K (1909) Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues Leipzig: Johann Ambrosius Barth. |
| [3] |
DeFelipe J, Alonso-Nanclares L, Arellano JI (2002) Microstructure of the neocortex: comparative aspects. J Neurocytol 31: 299-316. doi: 10.1023/A:1024130211265

|
| [4] |
Jacobs B, Scheibel AB (2002) Regional dendritic variation in primate cortical pyramidal cells. Cortical Areas: Unity and Diversity New York: CRC Press, 111-131. doi: 10.1201/9780203299296.pt2
|
| [5] |
Elston GN (2007) Specialization of the Neocortical Pyramidal Cell during Primate Evolution. Evolution of Nervous Systems, Volume IV Oxford: Academic Press, 191-242. doi: 10.1016/B0-12-370878-8/00164-6
|
| [6] |
Elston GN, Garey LJ (2013) The cytoarchitectonic map of Korbinian Brodmann: arealisation and circuit specialisation. Microstructural Parcellation of the Human Cerebral Cortex Berlin Heidelberg: Springer-Verlag, 3-32. doi: 10.1007/978-3-662-45766-5_1
|
| [7] |
Hafting T, Fyhn M, Molden S, et al. (2005) Microstructure of a spatial map in the entorhinal cortex. Nature 436: 801-806. doi: 10.1038/nature03721
|
| [8] |
Freiwald WA, Tsao DY (2010) Functional compartmentalization and viewpoint generalization within the macaque face-processing system. Science 330: 845-851. doi: 10.1126/science.1194908
|
| [9] |
DiCarlo JJ, Zoccolan D, Rust NC (2012) How does the brain solve visual object recognition? Neuron 73: 415-434. doi: 10.1016/j.neuron.2012.01.010
|
| [10] |
Kriegeskorte N, Kievit RA (2013) Representational geometry: integrating cognition, computation, and the brain. Trends Cognit Sci 17: 401-412. doi: 10.1016/j.tics.2013.06.007
|
| [11] |
Constantinescu AO, O'Reilly JX, Behrens TE (2016) Organizing conceptual knowledge in humans with a gridlike code. Science 352: 1464-1468. doi: 10.1126/science.aaf0941
|
| [12] |
Fusi S, Miller EK, Rigotti M (2016) Why neurons mix: high dimensionality for higher cognition. Curr Opin Neurobiol 37: 66-74. doi: 10.1016/j.conb.2016.01.010
|
| [13] |
Rolls ET (2016) Cerebral Cortex: Principles of Operation Oxford: Oxford University Press. doi: 10.1093/acprof:oso/9780198784852.001.0001
|
| [14] |
Chang L, Tsao DY (2017) The code for facial identity in the primate brain. Cell 169: 1013-1028. doi: 10.1016/j.cell.2017.05.011
|
| [15] |
Diedrichsen J, Kriegeskorte N (2017) Representational models: A common framework for understanding encoding, pattern-component, and representational-similarity analysis. PLOS Comput Biol 13: e1005508. doi: 10.1371/journal.pcbi.1005508
|
| [16] |
Landi SM, Freiwald WA (2017) Two areas for familiar face recognition in the primate brain. Science 357: 591-595. doi: 10.1126/science.aan1139
|
| [17] |
Kriegeskorte N, Diedrichsen J (2019) Peeling the onion of brain representations. Annu Rev Neurosci 42: 407-432. doi: 10.1146/annurev-neuro-080317-061906
|
| [18] |
Hinton GE (1989) Connectionist learning procedures. Artif Intell 40: 185-234. doi: 10.1016/0004-3702(89)90049-0
|
| [19] |
Schmidhuber J (2015) Deep learning in neural networks: An overview. Neural Netw 61: 85-117. doi: 10.1016/j.neunet.2014.09.003
|
| [20] |
Marblestone AH, Wayne G, Kording KP (2016) Toward an integration of deep learning and neuroscience. Front Comput Neurosci 10: 94. doi: 10.3389/fncom.2016.00094
|
| [21] |
Yamins DL, DiCarlo JJ (2016) Using goal-driven deep learning models to understand sensory cortex. Nat Neurosci 19: 356-365. doi: 10.1038/nn.4244
|
| [22] |
Beyeler M, Rounds EL, Carlson KD, et al. (2019) Neural correlates of sparse coding and dimensionality reduction. PLOS Comput Biol 15: e1006908. doi: 10.1371/journal.pcbi.1006908
|
| [23] |
Yang GR, Joglekar MR, Song HF, et al. (2019) Task representations in neural networks trained to perform many cognitive tasks. Nat Neurosci 22: 297-306. doi: 10.1038/s41593-018-0310-2
|
| [24] |
Ahissar M, Hochstein S (1993) Attentional control of early perceptual learning. Proc Natl Acad Sci USA 90: 5718-5722. doi: 10.1073/pnas.90.12.5718
|
| [25] | Adelson EH (2000) Lightness Perception and Lightness Illusions. The New Cognitive Neurosciences Cambridge: The MIT Press, 339-351. |
| [26] |
Fahle M (2004) Perceptual learning: A case for early selection. J Vis 4: 879-890. doi: 10.1167/4.10.4
|
| [27] |
Song Y, Ding Y, Fan S, et al. (2005) Neural substrates of visual perceptual learning of simple and complex stimuli. Clin Neurophysiol 116: 632-639. doi: 10.1016/j.clinph.2004.09.019
|
| [28] |
Sasaki Y, Nanez JE, Watanabe T (2010) Advances in visual perceptual learning and plasticity. Nat Rev Neurosci 11: 53-60. doi: 10.1038/nrn2737
|
| [29] |
Koster-Hale J, Saxe R (2013) Theory of mind: a neural prediction problem. Neuron 79: 836-848. doi: 10.1016/j.neuron.2013.08.020
|
| [30] |
Larcombe SJ, Kennard C, Bridge H (2017) Time course influences transfer of visual perceptual learning across spatial location. Vision Res 135: 26-33. doi: 10.1016/j.visres.2017.04.002
|
| [31] |
Buzsaki G, Moser EI (2013) Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat Neurosci 16: 130-138. doi: 10.1038/nn.3304
|
| [32] |
Rigotti M, Barak O, Warden MR, et al. (2013) The importance of mixed selectivity in complex cognitive tasks. Nature 497: 585-590. doi: 10.1038/nature12160
|
| [33] |
Moser MB, Rowland DC, Moser EI (2015) Place cells, grid cells, and memory. Cold Spring Harb Perspect Biol 7: a021808. doi: 10.1101/cshperspect.a021808
|
| [34] |
Saxena S, Cunningham JP (2019) Towards the neural population doctrine. Curr Opin Neurobiol 55: 103-111. doi: 10.1016/j.conb.2019.02.002
|
| [35] |
Prasad S, Galetta SL (2011) Anatomy and physiology of the afferent visual system. Handbook of Clinical Neurology Elsevier, 3-19. doi: 10.1016/B978-0-444-52903-9.00007-8
|
| [36] |
De Sousa AA, Proulx MJ (2014) What can volumes reveal about human brain evolution? A framework for bridging behavioral, histometric, and volumetric perspectives. Front Neuroanat 8: 51. doi: 10.3389/fnana.2014.00051
|
| [37] |
Recanzone GH, Schreiner CE, Merzenich MM (1993) Plasticity in the frequency representation of primary auditory cortex following discrimination training in adult owl monkeys. J Neurosci 3: 87-103. doi: 10.1523/JNEUROSCI.13-01-00087.1993
|
| [38] |
Wang X, Merzenich MM (1995) Remodelling of hand representation in adult. Nature 378: 2. doi: 10.1038/378071a0
|
| [39] |
Wallis G, Rolls ET (1997) Invariant face and object recognition in the visual system. Prog Neurobiol 51: 167-194. doi: 10.1016/S0301-0082(96)00054-8
|
| [40] |
Yang Z, Purves D (2004) The statistical structure of natural light patterns determines perceived light intensity. Proc Natl Acad Sci USA 101: 8745-8750. doi: 10.1073/pnas.0402192101
|
| [41] |
Smithson HE (2005) Sensory, computational and cognitive components of human colour constancy. Philos Trans R Soc Lond B Biol Sci 360: 1329-1346. doi: 10.1098/rstb.2005.1633
|
| [42] |
Corney D, Lotto RB (2007) What are lightness illusions and why do we see them? PLOS Comput Biol 3: 1790-1800. doi: 10.1371/journal.pcbi.0030180
|
| [43] |
Garrigan P, Kellman PJ (2008) Perceptual learning depends on perceptual constancy. Proc Natl Acad Sci USA 105: 2248-2253. doi: 10.1073/pnas.0711878105
|
| [44] |
Werner A (2014) Spatial and temporal aspects of chromatic adaptation and their functional significance for colour constancy. Vision Res 104: 80-89. doi: 10.1016/j.visres.2014.10.005
|
| [45] |
Kravitz DJ, Saleem KS, Baker CI, et al. (2013) The ventral visual pathway: an expanded neural framework for the processing of object quality. Trends Cognit Sci 17: 26-49. doi: 10.1016/j.tics.2012.10.011
|
| [46] |
Cichy RM, Pantazis D, Oliva A (2014) Resolving human object recognition in space and time. Nat Neurosci 17: 455-462. doi: 10.1038/nn.3635
|
| [47] |
Geskin J, Behrmann M (2018) Congenital prosopagnosia without object agnosia? A literature review. Cognit Neuropsychol 35: 4-54. doi: 10.1080/02643294.2017.1392295
|
| [48] |
De Beeck HP, Pillet I, Ritchie JB (2019) Factors determining where category-selective areas emerge in visual cortex. Trends Cognit Sci 23: 784-797. doi: 10.1016/j.tics.2019.06.006
|
| [49] |
Kar K, Kubilius J, Schmidt K, et al. (2019) Evidence that recurrent circuits are critical to the ventral stream's execution of core object recognition behavior. Nat Neurosci 22: 974-983. doi: 10.1038/s41593-019-0392-5
|
| [50] |
Eichenbaum H, Cohen NJ (2014) Can we reconcile the declarative memory and spatial navigation views on hippocampal function? Neuron 83: 764-770. doi: 10.1016/j.neuron.2014.07.032
|
| [51] |
Rolls ET, Wirth S (2018) Spatial representations in the primate hippocampus, and their functions in memory and navigation. Prog Neurobiol 171: 90-113. doi: 10.1016/j.pneurobio.2018.09.004
|
| [52] |
Tsutsui KI, Sakata H, Naganuma T, et al. (2002) Neural correlates for perception of 3D surface orientation from texture gradient. Science 298: 409-412. doi: 10.1126/science.1074128
|
| [53] |
Matthews N, Meng X, Xu P, et al. (2003) A physiological theory of depth perception from vertical disparity. Vision Res 43: 85-99. doi: 10.1016/S0042-6989(02)00401-7
|
| [54] |
Liu Y, Vogels R, Orban GA (2004) Convergence of depth from texture and depth from disparity in macaque inferior temporal cortex. J Neurosci 24: 3795-3800. doi: 10.1523/JNEUROSCI.0150-04.2004
|
| [55] |
Slater A, Mattock A, Brown E (1990) Size constancy at birth: Newborn infants' responses to retinal and real size. J Exp Child Psychol 49: 314-322. doi: 10.1016/0022-0965(90)90061-C
|
| [56] |
Liu Z (1999) Perceptual learning in motion discrimination that generalizes across motion directions. Pro Natl Acad Sci USA 96: 14085-14087. doi: 10.1073/pnas.96.24.14085
|
| [57] |
Sikl R, Simecek M, Porubanova-Norquist M, et al. (2013) Vision after 53 years of blindness. i-Percept 4: 498-507. doi: 10.1068/i0611
|
| [58] |
Lui JH, Hansen DV, Kriegstein AR (2011) Development and evolution of the human neocortex. Cell 146: 18-36. doi: 10.1016/j.cell.2011.06.030
|
| [59] | Fowler AE (1991) How Early Phonological Development Might Set the Stage for Phoneme Awareness. Phonological Processes in Literacy: A Tribute to Isabelle Y. Liberman Hillsdale, NJ: Erlbaum, 97-117. |
| [60] |
Kuhl PK, Williams KA, Lacerda F, et al. (1992) Linguistic experience alters phonetic perception in infants by 6 months of age. Science 255: 606-608. doi: 10.1126/science.1736364
|
| [61] |
Cheour M, Ceponiene R, Lehtokoski A, et al. (1998) Development of language-specific phoneme representations in the infant brain. Nat Neurosci 1: 351-353. doi: 10.1038/1561
|
| [62] |
Cowey A, Rolls ET (1974) Human cortical magnification factor and its relation to visual acuity. Exp Brain Res 21: 447-454. doi: 10.1007/BF00237163
|
| [63] |
Shum J, Hermes D, Foster BL, et al. (2013) A brain area for visual numerals. J Neurosci 33: 6709-6715. doi: 10.1523/JNEUROSCI.4558-12.2013
|
| [64] |
Blumberg J, Kreiman G (2010) How cortical neurons help us see: visual recognition in the human brain. J Clin Invest 120: 3054-3063. doi: 10.1172/JCI42161
|
| [65] |
Rolls ET, Baylis GC (1986) Size and contrast have only small effects on the responses to faces of neurons in the cortex of the superior temporal sulcus of the monkey. Exp Brain Res 65: 38-48. doi: 10.1007/BF00243828
|
| [66] |
Sharma J, Angelucci A, Sur M (2000) Induction of visual orientation modules in auditory cortex. Nature 404: 841-847. doi: 10.1038/35009043
|
| [67] |
Von Melchner L, Pallas SL, Sur M (2000) Visual behaviour mediated by retinal projections directed to the auditory pathway. Nature 404: 871-876. doi: 10.1038/35009102
|
| [68] | Simon HA (1962) The architecture of complexity. Proc Am Philos Soc 106: 467-482. |
| [69] |
Meunier D, Lambiotte R, Bullmore ET (2010) Modular and hierarchically modular organization of brain networks. Front Neurosci 4: 200. doi: 10.3389/fnins.2010.00200
|
| [70] |
Muckli L, Naumer MJ, Singer W (2009) Bilateral visual field maps in a patient with only one hemisphere. Proc Natl Acad Sci USA 106: 13034-13039. doi: 10.1073/pnas.0809688106
|
| [71] |
Borst A, Helmstaedter M (2015) Common circuit design in fly and mammalian motion vision. Nat Neurosci 18: 1067-1076. doi: 10.1038/nn.4050
|
| [72] |
De Andres-Bragado L, Sprecher SG (2019) Mechanisms of vision in the fruit fly. Curr Opin Insect Sci 36: 25-32. doi: 10.1016/j.cois.2019.06.005
|
| [73] |
Lee YJ, Nordstrom K (2012) Higher-order motion sensitivity in fly visual circuits. Proc Natl Acad Sci USA 109: 8758-8763. doi: 10.1073/pnas.1203081109
|
| [74] |
Tanaka K (1996) Inferotemporal cortex and object vision. Annu Rev Neurosci 19: 109-139. doi: 10.1146/annurev.ne.19.030196.000545
|
| [75] |
Hung CP, Kreiman G, Poggio T, et al. (2005) Fast readout of object identity from macaque inferior temporal cortex. Science 310: 863-866. doi: 10.1126/science.1117593
|
| [76] |
Kiani R, Esteky H, Mirpour K, et al. (2007) Object category structure in response patterns of neuronal population in monkey inferior temporal cortex. J Neurophysiol 97: 4296-4309. doi: 10.1152/jn.00024.2007
|
| [77] |
Yamins DL, Hong H, Cadieu CF, et al. (2014) Performance-optimized hierarchical models predict neural responses in higher visual cortex. Proc Natl Acad Sci USA 111: 8619-8624. doi: 10.1073/pnas.1403112111
|
| [78] |
Clarke A, Tyler LK (2015) Understanding what we see: how we derive meaning from vision. Trends Cognit Sci 19: 677-687. doi: 10.1016/j.tics.2015.08.008
|
| [79] |
Kohn A (2007) Visual adaptation: physiology, mechanisms, and functional benefits. J Neurophysiol 97: 3155-3164. doi: 10.1152/jn.00086.2007
|
| [80] |
Ress D, Heeger DJ (2003) Neuronal correlates of perception in early visual cortex. Nat Neurosci 6: 414-420. doi: 10.1038/nn1024
|
| [81] |
Jancke D, Chavane F, Naaman S, et al. (2004) Imaging cortical correlates of illusion in early visual cortex. Nature 428: 423-426. doi: 10.1038/nature02396
|
| [82] | O'Keefe J, Nadel L (1978) The hippocampus as a cognitive map Oxford: Clarendon Press. |
| [83] |
Brown MW, Aggleton JP (2001) Recognition memory: what are the roles of the perirhinal cortex and hippocampus? Nat Rev Neurosci 2: 51-61. doi: 10.1038/35049064
|
| [84] |
Eichenbaum H (2004) Hippocampus: cognitive processes and neural representations that underlie declarative memory. Neuron 44: 109-120. doi: 10.1016/j.neuron.2004.08.028
|
| [85] |
Leutgeb S, Leutgeb JK, Barnes CA, et al. (2005) Independent codes for spatial and episodic memory in hippocampal neuronal ensembles. Science 309: 619-623. doi: 10.1126/science.1114037
|
| [86] |
Konkel A, Cohen NJ (2009) Relational memory and the hippocampus: representations and methods. Front Neurosci 3: 166-174. doi: 10.3389/neuro.01.023.2009
|
| [87] |
Bassett DS, Bullmore ED (2006) Small-world brain networks. Neuroscientist 12: 512-523. doi: 10.1177/1073858406293182
|
| [88] |
Kanwisher N, Wojciulik E (2000) Visual attention: insights from brain imaging. Nat Rev Neurosci 1: 91-100. doi: 10.1038/35039043
|
| [89] |
Ungerleider SK (2000) Mechanisms of visual attention in the human cortex. Annu Rev Neurosci 23: 315-341. doi: 10.1146/annurev.neuro.23.1.315
|
| [90] |
Itti L, Koch C (2001) Computational modelling of visual attention. Nat Rev Neurosci 2: 194-203. doi: 10.1038/35058500
|
| [91] |
Lamme VA (2003) Why visual attention and awareness are different. Trends Cognit Sci 7: 12-18. doi: 10.1016/S1364-6613(02)00013-X
|
| [92] |
Serences JT, Yantis S (2006) Selective visual attention and perceptual coherence. Trends Cognit Sci 10: 38-45. doi: 10.1016/j.tics.2005.11.008
|
| [93] |
Mack ML, Love BC, Preston AR (2016) Dynamic updating of hippocampal object representations reflects new conceptual knowledge. Proc Natl Acad Sci USA 113: 13203-13208. doi: 10.1073/pnas.1614048113
|
| [94] |
White BJ, Berg DJ, Kan JY, et al. (2017) Superior colliculus neurons encode a visual saliency map during free viewing of natural dynamic video. Nat Commun 8: 1-9. doi: 10.1038/s41467-016-0009-6
|
| [95] |
Rademaker RL, Chunharas C, Serences JT (2019) Coexisting representations of sensory and mnemonic information in human visual cortex. Nat Neurosci 22: 1336-1344. doi: 10.1038/s41593-019-0428-x
|
| [96] |
Pang R, Lansdell BJ, Fairhall AL (2016) Dimensionality reduction in neuroscience. Curr Biol 26: R656-R660. doi: 10.1016/j.cub.2016.05.029
|
| [97] | Gosmann J, Eliasmith C (2020) CUE: A unified spiking neuron model of short-term and long-term memory. Psychol Rev Available from: https://doi.org/10.1037/rev0000250. |
| [98] |
Vyas S, Golub MD, Sussillo D, et al. (2020) Computation Through Neural Population Dynamics. Annu Rev Neurosci 43: 249-275. doi: 10.1146/annurev-neuro-092619-094115
|
| [99] |
Leskovec J, Sosic R (2016) Snap: A general-purpose network analysis and graph-mining library. ACM Trans Intell Syst Technol 8: 1-20. doi: 10.1145/2898361
|
| [100] |
Betzel RF, Bassett DS (2017) Generative models for network neuroscience: prospects and promise. J R Soc Interface 14: 20170623. doi: 10.1098/rsif.2017.0623
|
| [101] |
Guest O, Love BC (2017) What the success of brain imaging implies about the neural code. Elife 6: e21397. doi: 10.7554/eLife.21397
|
| [102] |
Avena-Koenigsberger A, Misic B, Sporns O (2018) Communication dynamics in complex brain networks. Nat Rev Neurosci 19: 17-33. doi: 10.1038/nrn.2017.149
|
| [103] |
Koutrouli M, Karatzas E, Paez-Espino D, et al. (2020) A Guide to Conquer the Biological Network Era Using Graph Theory. Front Bioeng Biotechnol 8: 34. doi: 10.3389/fbioe.2020.00034
|
| [104] |
Kriegeskorte N (2015) Deep neural networks: a new framework for modeling biological vision and brain information processing. Annu Rev Vis Sci 1: 417-446. doi: 10.1146/annurev-vision-082114-035447
|
| [105] |
Kell AJ, Yamins DL, Shook EN, et al. (2018) A task-optimized neural network replicates human auditory behavior, predicts brain responses, and reveals a cortical processing hierarchy. Neuron 98: 630-644. doi: 10.1016/j.neuron.2018.03.044
|
| [106] |
Bashivan P, Kar K, DiCarlo JJ (2019) Neural population control via deep image synthesis. Science 364: eaav9436. doi: 10.1126/science.aav9436
|
| [107] |
Stringer C, Pachitariu M, Steinmetz N, et al. (2019) High-dimensional geometry of population responses in visual cortex. Nature 571: 361-365. doi: 10.1038/s41586-019-1346-5
|
| [108] |
Atick JJ, Redlich AN (1990) Towards a theory of early visual processing. Neural Comput 2: 308-320. doi: 10.1162/neco.1990.2.3.308
|
| [109] |
Dan Y, Atick JJ, Reid RC (1996) Efficient coding of natural scenes in the lateral geniculate nucleus: experimental test of a computational theory. J Neurosci 16: 3351-3362. doi: 10.1523/JNEUROSCI.16-10-03351.1996
|
| [110] |
Doi E, Gauthier JL, Field GD, et al. (2012) Efficient coding of spatial information in the primate retina. J Neurosci 32: 16256-16264. doi: 10.1523/JNEUROSCI.4036-12.2012
|
| [111] |
Sporns O (2011) The non-random brain: efficiency, economy, and complex dynamics. Front Comput Neurosci 5: 5. doi: 10.3389/fncom.2011.00005
|
| [112] |
Smith EH, Horga G, Yates MJ, et al. (2019) Widespread temporal coding of cognitive control in the human prefrontal cortex. Nat Neurosci 22: 1883-1891. doi: 10.1038/s41593-019-0494-0
|
| [113] |
Yates AJ (1963) Delayed auditory feedback. Psychol Bull 60: 213-232. doi: 10.1037/h0044155
|
| [114] |
Marin OS, Perry DW (1999) Neurological aspects of music perception and performance. The psychology of music Academic Press, 653-724. doi: 10.1016/B978-012213564-4/50018-4
|
| [115] |
Barnes GR (2008) Cognitive processes involved in smooth pursuit eye movements. Brain Cogn 68: 309-326. doi: 10.1016/j.bandc.2008.08.020
|
| [116] |
David N, Newen A, Vogeley K (2008) The “sense of agency” and its underlying cognitive and neural mechanisms. Conscious Cognit 17: 523-534. doi: 10.1016/j.concog.2008.03.004
|
| [117] |
Rohrmeier MA, Koelsch S (2012) Predictive information processing in music cognition. A critical review. Int J Psychophysiol 83: 164-175. doi: 10.1016/j.ijpsycho.2011.12.010
|
| [118] |
Rohde M, Van Dam LC, Ernst MO (2014) Predictability is necessary for closed-loop visual feedback delay adaptation. J Vision 14: 4. doi: 10.1167/14.3.4
|
| [119] |
Okada K, Matchin W, Hickok G (2018) Neural evidence for predictive coding in auditory cortex during speech production. Psychon Bull Rev 25: 423-430. doi: 10.3758/s13423-017-1284-x
|
| [120] |
Lyu B, Choi HS, Marslen-Wilson WD, et al. (2019) Neural dynamics of semantic composition. Proc Natl Acad Sci USA 116: 21318-21327. doi: 10.1073/pnas.1903402116
|
| [121] |
Libet B, Gleason CA, Wright EW, et al. (1993) Time of conscious intention to act in relation to onset of cerebral activity (readiness-potential). Neurophysiology of Consciousness Boston: Birkhäuser, 249-268. doi: 10.1007/978-1-4612-0355-1_15
|
| [122] |
Haggard P, Clark S (2003) Intentional action: Conscious experience and neural prediction. Conscious Cogn 12: 695-707. doi: 10.1016/S1053-8100(03)00052-7
|
| [123] |
Haggard P (2005) Conscious intention and motor cognition. Trends Cognit Sci 9: 290-295. doi: 10.1016/j.tics.2005.04.012
|
| [124] |
Moore JW, Wegner DM, Haggard P (2009) Modulating the sense of agency with external cues. Conscious Cogn 18: 1056-1064. doi: 10.1016/j.concog.2009.05.004
|
| [125] |
Moore JW, Obhi SS (2012) Intentional binding and the sense of agency: a review. Conscious Cogn 21: 546-561. doi: 10.1016/j.concog.2011.12.002
|
| [126] |
Wen W, Yamashita A, Asama H (2015) The influence of action-outcome delay and arousal on sense of agency and the intentional binding effect. Conscious Cogn 36: 87-95. doi: 10.1016/j.concog.2015.06.004
|
| [127] |
Haggard P (2017) Sense of agency in the human brain. Nat Rev Neurosci 18: 196-207. doi: 10.1038/nrn.2017.14
|
| [128] |
Wegner DM (2002) The illusion of conscious will The MIT press. doi: 10.7551/mitpress/3650.001.0001
|
| [129] |
Komer B, Eliasmith C (2016) A unified theoretical approach for biological cognition and learning. Curr Opin Behav Sci 11: 14-20. doi: 10.1016/j.cobeha.2016.03.006
|
| [130] |
Bullmore E, Sporns O (2012) The economy of brain network organization. Nat Rev Neurosci 13: 336-349. doi: 10.1038/nrn3214
|
| [131] |
Stiles J, Jernigan TL (2010) The basics of brain development. Neuropsychol Rev 20: 327-348. doi: 10.1007/s11065-010-9148-4
|
| [132] |
Budday S, Steinmann P, Kuhl E (2015) Physical biology of human brain development. Front Cell Neurosci 9: 257. doi: 10.3389/fncel.2015.00257
|
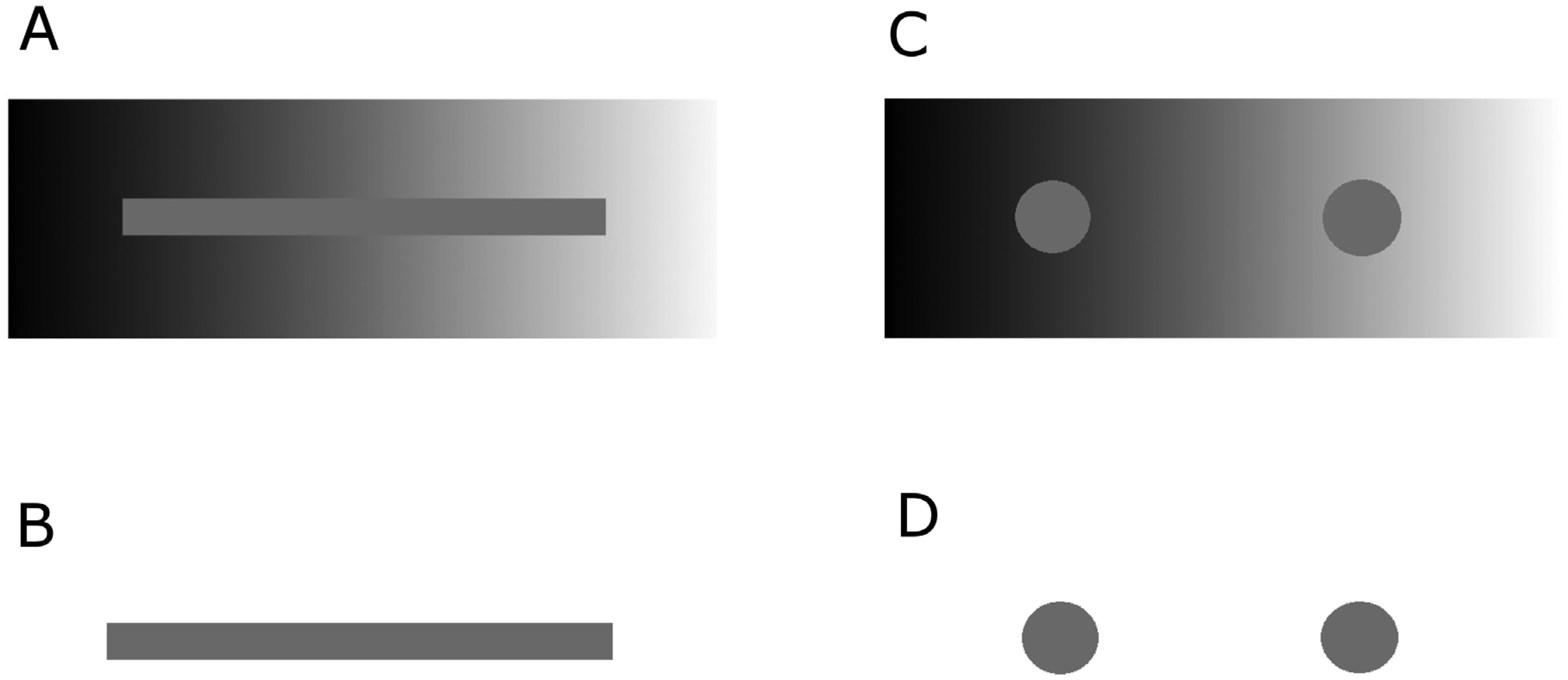
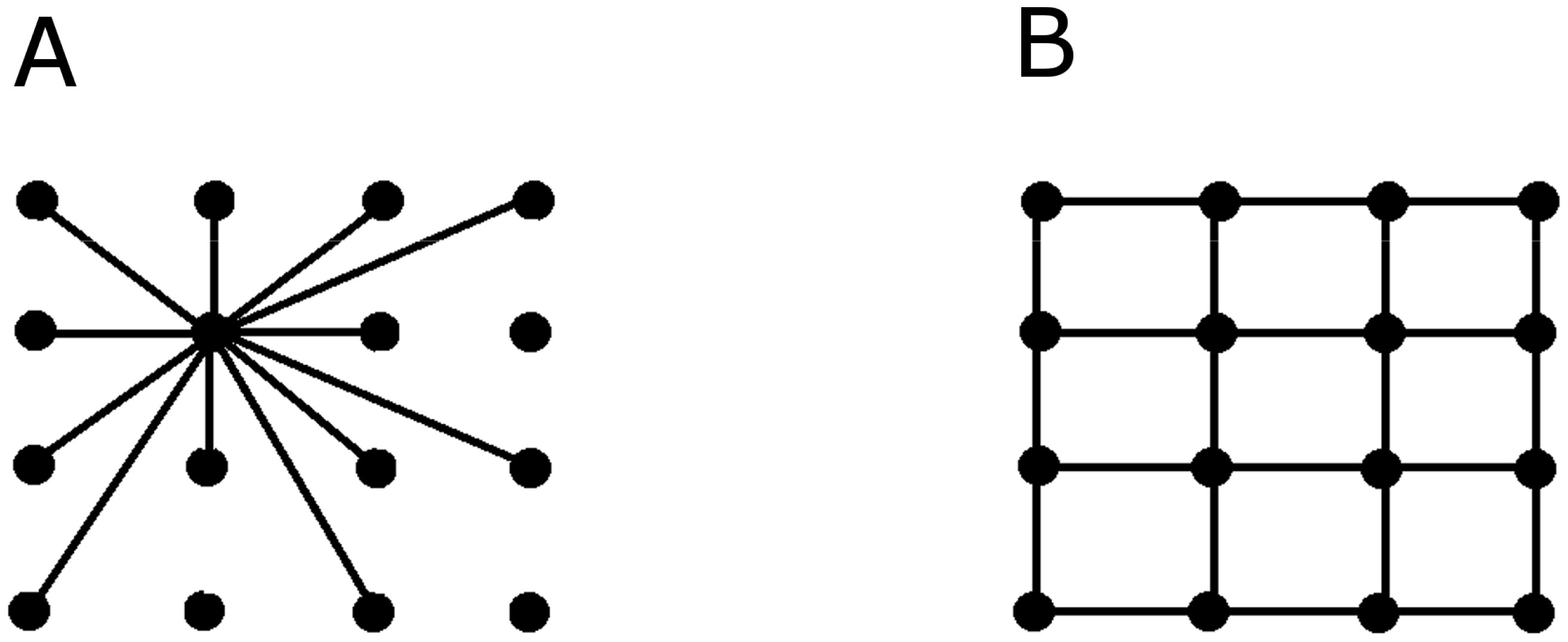
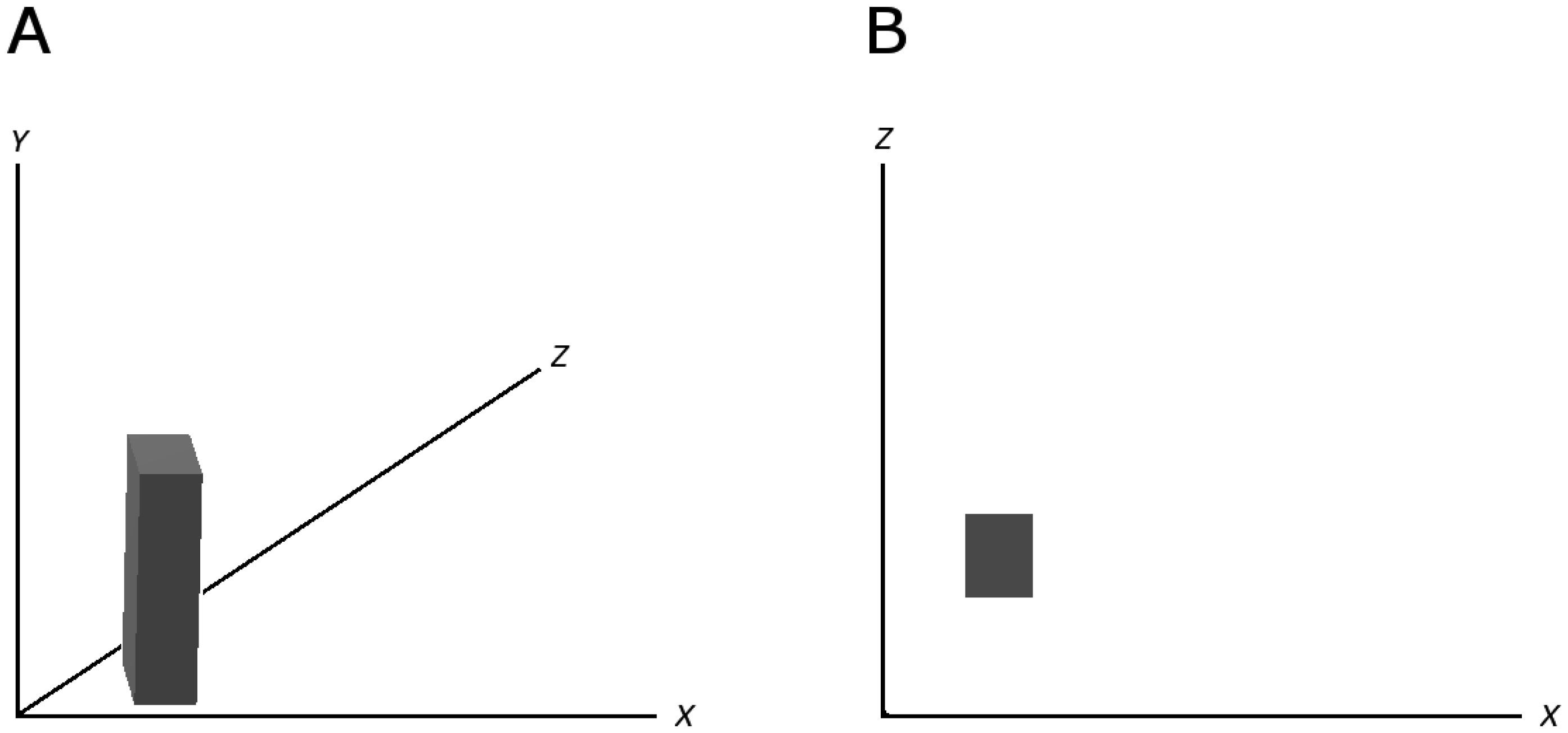
Figures(3)

Robert Friedman. Themes of advanced information processing in the primate brain[J]. AIMS Neuroscience, 2020, 7(4): 373-388. doi: 10.3934/Neuroscience.2020023







 DownLoad:
DownLoad: