Citation: Wei-Bin Zhang. Growth, environmental change and business cycles in a multi-regional economy[J]. Green Finance, 2019, 1(2): 205-220. doi: 10.3934/GF.2019.2.205

| [1] |
Antonakakis N, Chatziantoniou I, Filis G (2017) Energy Consumption, CO2 Emissions, and Economic Growth: An Ethical Dilemma. Renew Sust Energy Rev 68: 808–824. doi: 10.1016/j.rser.2016.09.105

|
| [2] |
Apergis N, Payne JE (2014) Renewable Energy, Output, CO2 Emissions, and Fossil Fuel Prices in Central America: Evidence from a Nonlinear Panel Smooth Transition Vector Error Correction Model. Energy Econ 42: 226–232. doi: 10.1016/j.eneco.2014.01.003
|
| [3] | Chen Y, Irwin EG, Jayaprakash C (2013) Population Dispersion vs. Concentration in a Two-Region Migration Model with Endogenous Natural Amenities. J Reg Sci 53: 256–273. |
| [4] | Chiarella C, Flaschel P (2000) The Dynamics of Keynesian Monetary Growth: Macro Foundations, Cambridge University Press, Cambridge. |
| [5] |
Debarsy N, Ertur C (2019) Interaction Matrix Selection in Spatial Autoregressive Models with an Application to Growth Theory. Reg Sci Urban Econ 75: 49–69. doi: 10.1016/j.regsciurbeco.2019.01.002
|
| [6] | Diamond DB, Tolley GS (1981) The Economics of Urban Amenities, New York: Academic Press. |
| [7] |
Forslid R, Ottaviano GIP (2003) Trade and Location: Two Analytically Solvable Models. J Econ Geogr 3: 229–240. doi: 10.1093/jeg/3.3.229
|
| [8] | Fujita M, Krugman P, Venables A (1999) The Spatial Economy, Cambridge, MA: MIT Press. |
| [9] |
Fullerton D, Kim SR (2008) Environmental investment and policy with distortionary taxes, and endogenous growth. J Environ Econ Manage 56: 141–154. doi: 10.1016/j.jeem.2008.02.001
|
| [10] | Gandolfo G (2005) Economic Dynamics, Berlin: Springer. |
| [11] |
Gassebner M, Lamlay MJ, Sturmz JE (2011) Determinants of Pollution: What Do We Really Know? Oxford Econ Pap 63: 568–595. doi: 10.1093/oep/gpq029
|
| [12] |
Glaeser EL, Maré DC (2001) Cities and Skills. J Labor Econ 19: 316–342. doi: 10.1086/319563
|
| [13] | Hirschman AO (1958) The Strategy of Economic Development, New Haven: Yale University Press. |
| [14] |
Krugman P (1991) Increasing Returns and Economic Geography. J Polit Econ 99: 483–499. doi: 10.1086/261763
|
| [15] |
Krugman P (1993) On the Number and Location of Cities. Eur Econ Rev 37: 293–298. doi: 10.1016/0014-2921(93)90017-5
|
| [16] | Kuznets S (1966) Modern Economic Growth, Rate Structure, and Spread, New Haven: Yale University Press. |
| [17] |
Lai YB (2019) Environmental Policy Competition and Heterogeneous Capital Endowments. Reg Sci Urban Econ 75: 107–119. doi: 10.1016/j.regsciurbeco.2019.01.011
|
| [18] |
Landry SM, Chakraborty J (2009) Street Trees and Equity: Evaluating the Spatial Distribution of an Urban Amenity. Environ Plann A 41: 2651–2670. doi: 10.1068/a41236
|
| [19] |
Lanz B, Provins A (2013) Valuing Local Environmental Amenity with Discrete Choice Experiments: Spatial Scope Sensitivity and Heterogeneous Marginal Utility of Income. Environ Resour Econ 56: 105–130. doi: 10.1007/s10640-013-9648-9
|
| [20] | Lin CYC, Liscow ZD (2012) Endogeneity in the Environmental Kuznets Curve: An Instrumental Variables Approach. Am J Agric Econ 95: 268–274. |
| [21] | Lorenz HW (1993) Nonlinear Dynamic Economics and Chaotic Motion, Springer-Verlag, Berlin. |
| [22] | Myrdal G (1957) Economic Theory and Underdeveloped Regions, London: Duckworth. |
| [23] |
Partridge MD, Rickman DS, Ali K, et al. (2008) Lost in Space: Population Dynamics in the American Hinterlands and Small Cities. J Econ Geogr 8: 727–757. doi: 10.1093/jeg/lbn038
|
| [24] |
Plouder GC (1972) A model of waste accumulation and disposal. Can J Econ 5: 119–125. doi: 10.2307/133997
|
| [25] | Puu T (2011) Nonlinear Economic Dynamics, Berlin: Springer. |
| [26] |
Saracoglu DS, Roe TL (2019) Regional Cost-of-Living Differentials, Rural-Urban Migration, and the Contribution to Economic Growth. Pap Reg Sci 98: 973–994. doi: 10.1111/pirs.12404
|
| [27] | Shone R (2002) Economic Dynamics-Phase Diagrams and Their Economic Application, Cambridge University Press, Cambridge. |
| [28] |
Tabuchi T (2014) Historical Trends of Agglomeration to the Capital Region and New Economic Geography. Reg Sci Urban Econ 44: 50–59. doi: 10.1016/j.regsciurbeco.2013.11.004
|
| [29] |
Tivadar M, Jayet H (2019) Endogenous Generation of Amenities and the Dynamics of City Structure. Reg Sci Urban Econ 74: 49–56. doi: 10.1016/j.regsciurbeco.2018.10.005
|
| [30] |
Yang Z, Zhao Y (2014) Energy Consumption, Carbon Emissions, and Economic Growth in India: Evidence from Directed Acyclic Graphs. Econ Model 38: 533–540. doi: 10.1016/j.econmod.2014.01.030
|
| [31] |
Walenta J (2018) The Limits to Private-Sector Climate Change Action: The Geographies of Corporate Climate Governance. Econ Geogr 94: 461–484. doi: 10.1080/00130095.2018.1474078
|
| [32] | Zhang WB (1991) Synergetic Economics, Springer-Verlag, Heidelberg. |
| [33] | Zhang WB (1993) Wages, Service Prices and Rent - Urban Division of Labor and Amenities. Seoul J Econ 6: 97–113. |
| [34] | Zhang WB (2005) Differential Equations, Bifurcations, and Chaos in Economics, Singapore: World Scientific. |
| [35] | Zhang WB (2006) Discrete Dynamical Systems, Bifurcations and Chaos in Economics, Elsevier: Amsterdam. |
| [36] |
Zhang WB (2016) A Study of Regional Environmental Change and Economic Growth by Extending Solow's Growth Model. Ekon Yaklasim 27: 223–254. doi: 10.5455/ey.35931
|
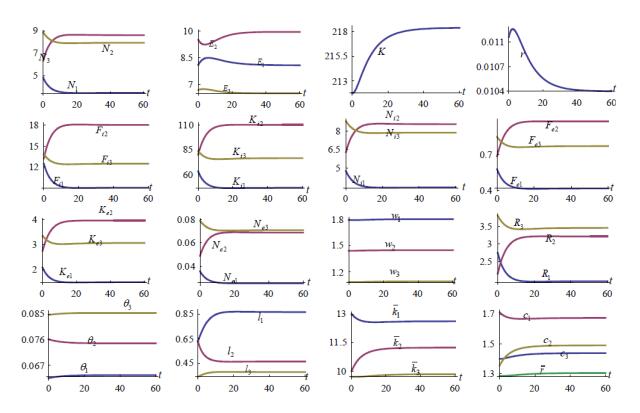
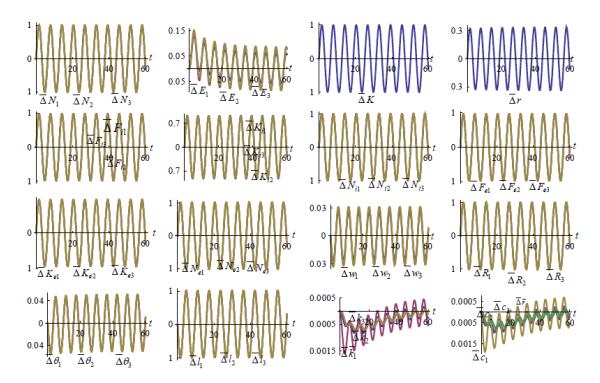
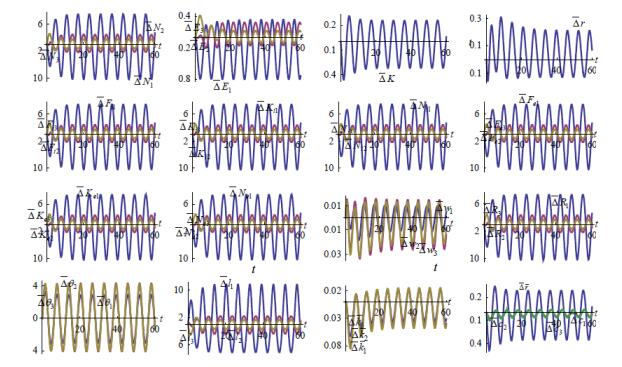
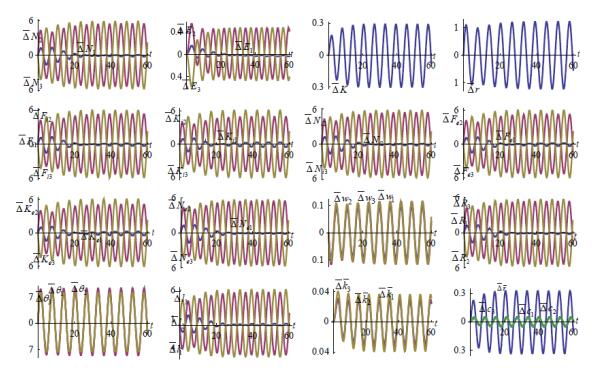
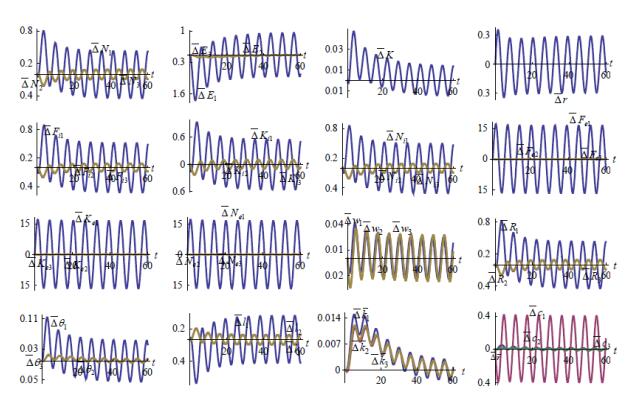
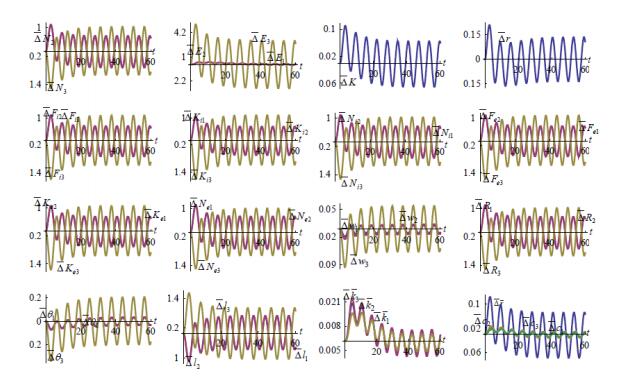
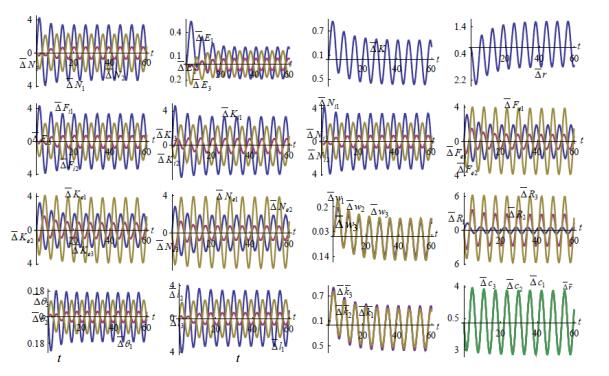
Figures(7)

Wei-Bin Zhang. Growth, environmental change and business cycles in a multi-regional economy[J]. Green Finance, 2019, 1(2): 205-220. doi: 10.3934/GF.2019.2.205







 DownLoad:
DownLoad: