Citation: Noor-Azira Abdul-Mutalib, Syafinaz Amin Nordin, Malina Osman, Ahmad Muhaimin Roslan, Natsumi Ishida, Kenji Sakai, Yukihiro Tashiro, Kosuke Tashiro, Toshinari Maeda, Yoshihito Shirai. The prevalence of foodborne pathogenic bacteria on cutting boards and their ecological correlation with background biota[J]. AIMS Microbiology, 2016, 2(2): 138-151. doi: 10.3934/microbiol.2016.2.138

| [1] |
Vongkamjan K, Wiedmann M (2015) Starting from the bench- Prevention and control of foodborne and zoonotic diseases. Prev Vet Med 118: 189–195. doi: 10.1016/j.prevetmed.2014.11.004

|
| [2] | Wang B, Wang Q, Cai Z, et al. (2015) Simultaneous, rapid and sensitive detection of three food-borne pathogenic bacteria using multicolor quantum dot probes based on multiplex fluoroimmunoassay in food samples. LWT - Food Sci Technol 61: 368–376. |
| [3] |
Larsen MH, Dalmasso M, Ingmer H, et al. (2014) Persistence of foodborne pathogens and their control in primary and secondary food production chains. Food Control 44: 92–109. doi: 10.1016/j.foodcont.2014.03.039
|
| [4] |
Kothary MH, Babu US (2001) Infective dose of foodborne pathogens in volunteers: A review. J Food Safety 21: 49–68. doi: 10.1111/j.1745-4565.2001.tb00307.x
|
| [5] | Tuttle J, Gomez T, Doyle MP, et al. (1999) Lessons from a large outbreak of Escherichia coli O157:H7 infections: Insights into the infectious dose and method of widespread contamination of hamburger patties. Epidemiol Infect 122: 185–192. |
| [6] |
Pina-Pérez MC, Silva-Angulo AB, Rodrigo D, et al. (2012) A preliminary exposure assessment model for Bacillus cereus cells in a milk based beverage: Evaluating high pressure processing and antimicrobial interventions. Food Control 26: 610–613. doi: 10.1016/j.foodcont.2012.01.063
|
| [7] | Fakruddin M, Rahaman MM, Ahmed MM, et al. (2013) Cronobacter sakazakii (Enterobacter sakazakii): An emerging food borne pathogen. Int J Biomed Adv Res 4: 349–359. |
| [8] | Sreedharan A, Jones C, Schneider K Preventing Foodborne Illness: Yersiniosis, 2012. Available from: http://edis.ifas.ufl.edu/fs193 |
| [9] |
Crost EH, Pujol A, Ladiré M, et al. (2010). Production of an antibacterial substance in the digestive tract involved in colonization-resistance against Clostridium perfringens. Anaerobe 16: 597–603. doi: 10.1016/j.anaerobe.2010.06.009
|
| [10] |
Kaiko GE, Stappenbeck TS (2014) Host-microbe interactions shaping the gastrointestinal environment. Trends Immunol 35: 538–548. doi: 10.1016/j.it.2014.08.002
|
| [11] |
Francis GA, O'Beirne D (1998). Effects of the indigenous microflora of minimally processed lettuce on the survival and growth of Listeria innocua. Int J Food Sci Tech 33: 477–488. doi: 10.1046/j.1365-2621.1998.00199.x
|
| [12] | Schiemann DA, Olson SA (1984). Antagonism by gram-negative bacteria to growth of Yersinia enterocolitica in mixed cultures. Appl Environ Microbiol 48: 539–544. |
| [13] |
Alegre I, Viñas I, Usall J, et al. (2013) Antagonistic effect of Pseudomonas graminis CPA-7 against foodborne pathogens in fresh-cut apples under simulated commercial conditions. Food Microbiol 33: 139–148. doi: 10.1016/j.fm.2012.09.007
|
| [14] | Ashraf R, Vasiljevic T, Smith SC, et al. (2014) Effect of cell-surface components and metabolites of lactic acid bacteria and probiotic organisms on cytokine production and induction of CD25 expression in human peripheral mononuclear cells. J Dairy Sci 97: 2542–2558. |
| [15] | O'Bryan CA, Crandall PG, Ricke SC, et al. (2015) Lactic acid bacteria (LAB) as antimicrobials in food products: Types and mechanisms of action. Handbook of Natural Antimicrobials for Food Safety and Quality 117–136. |
| [16] |
Pedras MS, Ismail N, Quail JW, et al. (2003) Structure, chemistry, and biological activity of pseudophomins A and B, new cyclic lipodepsipeptides isolated from the biocontrol bacterium Pseudomonas fluorescens. Phytochemistry 62: 1105–1114. doi: 10.1016/S0031-9422(02)00617-9
|
| [17] | Trotel-Aziz P, Couderchet M, Biagianti S, et al. (2008) Characterization of new bacterial biocontrol agents Acinetobacter, Bacillus, Pantoea and Pseudomonas spp. mediating grapevine resistance against Botrytis cinerea. Environ Exp Bot 64: 21–32. |
| [18] |
Joen YS, Chun J, Kim BS (2013) Identification of household bacterial community and analysis of species shared with human microbiome. Curr Microbiol 67: 557–563. doi: 10.1007/s00284-013-0401-y
|
| [19] |
Jung J, Lee SH, Jin HM, et al. (2014) Pyrosequencing-based analysis of bacterial community and metabolites profiles in Korean traditional seafood fermentation: A flatfish-fermented seafood. Biosci Biotech Bioch 78: 908–910. doi: 10.1080/09168451.2014.895659
|
| [20] |
Koyanagi T, Nakagawa A, Kiyohara M, et al. (2013) Pyrosequencing analysis of microbiota in Kaburazushi, a traditional medieval sushi in Japan. Biosci Biotech Bioch 77: 2125–2130. doi: 10.1271/bbb.130550
|
| [21] |
Nam YD, Yi SH, Lim SI (2012) Bacterial diversity of cheonggukjang, a traditional Korean fermented food, analyzed by barcoded pyrosequencing. Food Control 28: 135–142. doi: 10.1016/j.foodcont.2012.04.028
|
| [22] |
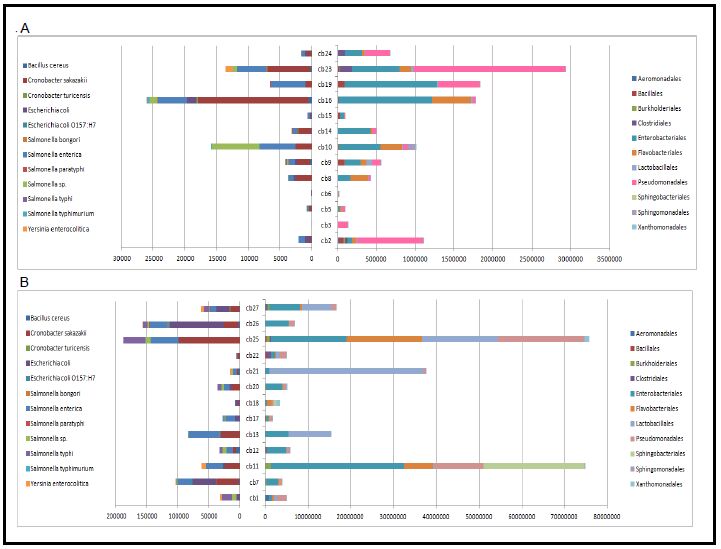
Abdul-Mutalib NA, Amin Nordin S, Osman M, et al. (2015) Pyrosequencing analysis of microbial community and food-borne bacteria on restaurant cutting boards collected in Seri Kembangan, Malaysia, and their correlation with grades of food premises. Int J Food Microbiol 200: 57–65. doi: 10.1016/j.ijfoodmicro.2015.01.022
|
| [23] |
Gómez D, Ariño A, Carramiñana JJ, et al. (2012) Sponge versus mini-roller for the surface microbiological control of Listeria monocytogenes, total aerobic mesophiles and Enterobacteriaceae in the meat industry. Food Control 27: 242–247. doi: 10.1016/j.foodcont.2012.03.031
|
| [24] |
Kakizaki E, Ogura Y, Kozawa S, et al. (2012) Detection of diverse aquatic microbes in blood and organs of drowning victims: First metagenomic approach using high-throughput 454-pyrosequencing. Forensic Sci Int 220: 135–146. doi: 10.1016/j.forsciint.2012.02.010
|
| [25] |
Hund A, Dzieciol M, Schmitz-Esser S, et al. (2015) Characterization of mucosa-associated bacterial communities in abomasal ulcers by pyrosequencing. Vet Microbiol 177:132–141. doi: 10.1016/j.vetmic.2015.02.023
|
| [26] |
McCann CD, Moore MS, May LS, et al. (2015) Evaluation of real-time PCR and pyrosequencing for screening incubating blood culture bottles from adults with suspected bloodstream infection. Diagn Microbiol Infect Dis 81: 158–162. doi: 10.1016/j.diagmicrobio.2014.11.014
|
| [27] | Prest EI, El-Chakhtoura J, Hammes F, et al. (2014) Combining flow cytometry and 16S rRNA gene pyrosequencing: A promising approach for drinking water monitoring and characterization. Water Res 63: 179–189. |
| [28] |
Jay JM (1996) Microorganisms in fresh ground meats: The relative safety of products with low versus high numbers. Meat Sci 43: 59–66. doi: 10.1016/0309-1740(96)00055-1
|
| [29] | Shoaie S, Karlsson F, Mardinoglu A, et al. (2013) Understanding the interactions between bacteria in the human gut through metabolic modelling. Sci Rep 3: 2532. |
| [30] |
Vogt SL, Pena-Díaz J, Finlay BB (2015) Chemical communication in the gut: Effects of microbiota-generated metabolites on gastrointestinal bacterial pathogens. Anaerobe 34: 106–115 doi: 10.1016/j.anaerobe.2015.05.002
|
| [31] |
Abell GCJ, McOrist AL (2007) Assessment of the diversity and stability of faecal bacteria from healthy adults using molecular methods. Microb Ecol Health Dis 19: 229–240. doi: 10.1080/08910600701662248
|
| [32] | Rai R, Saraswat VA, Dhiman RK (2015) Gut microbiota: Its role in hepatic encephalopathy. J Clin Exp Hepatol 5: S29–S35 |
| [33] |
Thiennimitr P, Winter SE, Bäumler AJ (2012) Salmonella, the host and its microbiota. Curr Opin Microbiol 15:108–114. doi: 10.1016/j.mib.2011.10.002
|
| [34] |
Paredes-Sabja D, Setlow P, Sarker MR (2011) Germination of spores of Bacillales and Clostridiales species: mechanisms and proteins involved. Trends Microbiol 19: 85–94. doi: 10.1016/j.tim.2010.10.004
|
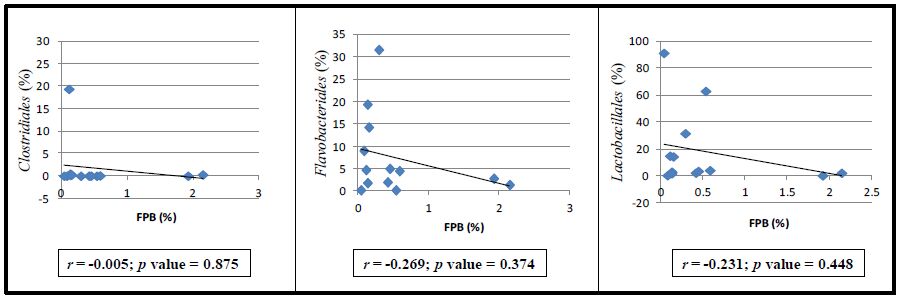
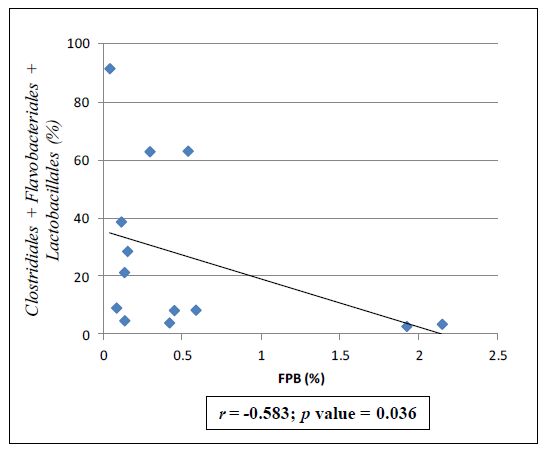
| [35] | Ferrario C, Taverniti V, Milani C, et al. (2014) Modulation of fecal Clostridiales bacteria and butyrate by probiotic intervention with Lactobacillus paracasei DG varies among healthy adults. J Nutr 144: 1787–1796. |
| [36] |
Baumgart M, Dogan B, Rishniw M, et al. (2007) Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn's disease involving the ileum. ISME J 1: 403–418. doi: 10.1038/ismej.2007.52
|
| [37] |
Kamada N, Chen GY, Inohara N, et al. (2013) Control of pathogens and pathobionts by the gut microbiota. Nat Immunol 14: 685–690. doi: 10.1038/ni.2608
|
| [38] | Stecher B, Hardt WD (2011) Mechanisms controlling pathogen colonization of the gut. Curr Opin Microbiol 14: 82–91. |
| [39] |
Vincent C, Stephens DA, Loo VG, et al. (2013) Reductions in intestinal Clostridiales precede the development of nosocomial Clostridium difficile infection. Microbiome 1: 18. doi: 10.1186/2049-2618-1-18
|
| [40] |
Schommer NN, Gallo RL (2013) Structure and function of the human skin microbiome. Trends Microbiol 21: 660–668. doi: 10.1016/j.tim.2013.10.001
|
| [41] | Kolton M, Sela N, Elad Y, et al. (2013) Comparative genomic analysis indicates that niche adaptation of terrestrial Flavobacteria is strongly linked to plant glycan metabolism. PLoS ONE 8: e76704. |
| [42] |
Chen HL, Lu JH, Wang HH, et al. (2014) Clinical analysis of Enterobacter bacteremia in pediatric patients: A 10-year study. J Microbiol Immunol Infect 47: 381–386. doi: 10.1016/j.jmii.2013.03.016
|
| [43] | Schlaeppi K, Dombrowski N, van Themaat RGOEVL, et al. (2013) Quantitative divergence of the bacterial root microbiota in Arabidopsis thaliana relatives. Proc Nat Acad Sci 111: 585–592. |
| [44] |
Grossart HP, Schlingloff A, Bernhard M, et al. (2004) Antagonistic activity of bacteria isolated from organic aggregates of the German Wadden Sea. FEMS Microbiol Ecol 47: 387–396. doi: 10.1016/S0168-6496(03)00305-2
|
| [45] | IuD S, Kruglikova LF, Vasiuk LF, et al. (1996) A new metabolite with fungistatic and bacteriostatic activity, produced by strain L-30 of Flavobacterium sp. Antibiot Khimioter 41: 6–12. |
| [46] | Tyc O, van den Berg M, Gerards S, et al. (2014) Impact of interspecific interactions on antimicrobial activity among soil bacteria. Front Microbiol 5: 567. |
| [47] | Goecke F, Labes A, Wiese J, et al. (2013) Phylogenetic analysis and antibiotic activity of bacteria isolated from the surface of two co-occurring macroalgae from the Baltic Sea. Eur J Phycol 8: 47–60. |
| [48] |
Salvetti E, Fondi M, Fani R, et al. (2013) Evolution of lactic acid bacteria in the order Lactobacillales as depicted by analysis of glycolysis and pentose phosphate pathways. Syst Appl Microbiol 36: 291–305. doi: 10.1016/j.syapm.2013.03.009
|
| [49] |
de Almeida Júnior WLG, Ferrari IS, de Souza JV, et al (2015). Principal criteria for selection of lactic acid bacteria for potential use as probiotic in foods. Afr J Microbiol Res 9: 671–686. doi: 10.5897/AJMR2014.7226
|
| [50] | Ballester JM, Ballester M, Belaich JP, (1980) Purification of the viridicin produced by Aerococcus viridians. Antimicrob Agents Ch 17: 784–788. |
| [51] | Papagianni M, Anastasiadou S (2009) Pediocins: The bacteriocins of Pediococci. Sources, production, properties and applications. Microbial Cell Factories 8: 3. |
| [52] |
Alegría A1, Delgado S, Roces C, et al. (2010) Bacteriocins produced by wild Lactococcus lactis strains isolated from traditional, starter-free cheeses made of raw milk. Int J Food Microbiol 143: 61–66. doi: 10.1016/j.ijfoodmicro.2010.07.029
|
| [53] |
Dogsa I, Choudhary KS, Marsetic Z, et al. (2014) ComQXPA quorum sensing systems may not be unique to Bacillus subtilis: A census in prokaryotic genomes. PLoS ONE 9: e96122. doi: 10.1371/journal.pone.0096122
|
| [54] |
Pantaléon V, Bouttier S, Soavelomandroso AP, et al. (2014) Biofilms of Clostridium species. Anaerobe 30: 193–198. doi: 10.1016/j.anaerobe.2014.09.010
|
| [55] |
Wang WZ, Morohoshi T, Someya N, et al. (2012) Aidc, a novel N-Acylhomoserine Lactonase from the potato root-associated Cytophaga-Flavobacteria-Bacteroides (CFB) group bacterium Chryseobacterium sp. strain StRB126. Appl Environ Microb 78: 7985–7992. doi: 10.1128/AEM.02188-12
|
| [56] |
LaSarre B, Federle MJ (2013) Exploiting quorum sensing to confuse bacterial pathogens. Microbiol Mol Biol Rev 77: 73–111. doi: 10.1128/MMBR.00046-12
|
| [57] | Macwana S, Muriana PM (2012) Spontaneous bacteriocin resistance in Listeria monocytogenes as a susceptibility screen for identifying different mechanisms of resistance and modes of action by bacteriocins of lactic acid bacteria J Microbiol Meth 88: 7–13. |
| [58] | Vermeulen A, Devlieghere F, Bernaerts K, et al. (2007) Growth/no growth models describing the influence of pH, lactic and acetic acid on lactic acid bacteria developed to determine the stability of acidified sauces. Int J Food Microbiol 119: 258–269. |
| [59] | Ghanbari M, Jami M, Kneifel W, et al. (2013) Antimicrobial activity and partial characterization of bacteriocins produced by lactobacilli isolated from Sturgeon fish. Food Control 32: 379–385. |
| [60] |
Hwanhlem N, Chobert JM, H-Kittikun A (2014) Bacteriocin-producing lactic acid bacteria isolated from mangrove forests in southern Thailand as potential bio-control agents in food: Isolation, screening and optimization. Food Control 41: 202–211. doi: 10.1016/j.foodcont.2014.01.021
|
| [61] | da Silva Sabo S, Vitolo M, González JMD, et al. (2014) Overview of Lactobacillus plantarum as a promising bacteriocin producer among lactic acid bacteria. Food Res Int 64: 527–536. |
| [62] |
Leong KH, Chen YS, Lin YH, et al. (2013) Weissellicin L, a novel bacteriocin from sian-sianzih-isolated Weissella hellenica 4-7. J Appl Microbiol 115: 70–76. doi: 10.1111/jam.12218
|
Figures(3) / Tables(4)

Noor-Azira Abdul-Mutalib, Syafinaz Amin Nordin, Malina Osman, Ahmad Muhaimin Roslan, Natsumi Ishida, Kenji Sakai, Yukihiro Tashiro, Kosuke Tashiro, Toshinari Maeda, Yoshihito Shirai. The prevalence of foodborne pathogenic bacteria on cutting boards and their ecological correlation with background biota[J]. AIMS Microbiology, 2016, 2(2): 138-151. doi: 10.3934/microbiol.2016.2.138







 DownLoad:
DownLoad: