Citation: Wilfredo Lopez, Alexis M. Page, Darby J. Carlson, Brad L. Ericson, Matyas F. Cserhati, Chittibabu Guda, Kimberly A. Carlson. Analysis of immune-related genes during Nora virus infection of Drosophila melanogaster using next generation sequencing[J]. AIMS Microbiology, 2018, 4(1): 123-139. doi: 10.3934/microbiol.2018.1.123

| [1] |
Habayeb MS, Ekengren SK, Hultmark D (2006) Nora virus, a persistent virus in Drosophila, defines a new picorna-like virus family. J Gen Virol 87: 3045–3051. doi: 10.1099/vir.0.81997-0

|
| [2] |
Webster CL, Waldron FM, Robertson S, et al. (2015) The discovery, distribution, and evolution of viruses associated with Drosophila melanogaster. PLoS Biol 13: e1002210. doi: 10.1371/journal.pbio.1002210
|
| [3] |
Johnson KN, Christian PD (1998) The novel genome organization of the insect picorna-like virus Drosophila C virus suggests this virus belongs to a previously undescribed virus family. J Gen Virol 79: 191–203. doi: 10.1099/0022-1317-79-1-191
|
| [4] |
Habayeb MS, Jens-Ola E, Dan H (2009) Nora virus persistent infections are not affected by the RNAi machinery. PLoS One 4: e5731. doi: 10.1371/journal.pone.0005731
|
| [5] | Ericson BL, Carlson DJ, Carlson KA (2016) Characterization of Nora virus structural proteins via Western blot analysis. Scientifica 2016: 9067848. |
| [6] |
Sadanandan SA, Ekström JO, Jonna VR, et al. (2016) VP3 is crucial for the stability of Nora virus virions. Virus Res 223: 20–27. doi: 10.1016/j.virusres.2016.06.011
|
| [7] |
Ekström JO, Habayeb MS, Srivastava V, et al. (2011) Drosophila Nora virus capsid proteins differ from those of other picorna-like viruses. Virus Res 160: 51–58. doi: 10.1016/j.virusres.2011.05.006
|
| [8] |
Habayeb MS, Cantera R, Casanova G, et al. (2009) The Drosophila Nora virus is an enteric virus, transmitted via feces. J Invertebr Pathol 101: 29–33. doi: 10.1016/j.jip.2009.02.003
|
| [9] |
De Gregorio E, Spellman PT, Tzou P, et al. (2002) The Toll and Imd pathways are the major regulators of the immune response in Drosophila. EMBO J 21: 2568–2579. doi: 10.1093/emboj/21.11.2568
|
| [10] |
Ferreira AG, Naylor H, Esteves SS, et al. (2014) The Toll-dorsal pathway is required for resistance to viral oral infection in Drosophila. PLoS Pathog 10: e1004507. doi: 10.1371/journal.ppat.1004507
|
| [11] |
Baeg GH, Zhou R, Perrimon N (2005) Genome-wide RNAi analysis of JAK/STAT signaling components in Drosophila. Gene Dev 19: 1861–1870. doi: 10.1101/gad.1320705
|
| [12] |
Zambon RA, Vakharia VN, Wu LP (2006) RNAi is an antiviral immune response against a dsRNA virus in Drosophila melanogaster. Cell Microbiol 8: 880–889. doi: 10.1111/j.1462-5822.2006.00688.x
|
| [13] |
Cordes EJ, Licking-Murray KD, Carlson KA (2013) Differential gene expression related to Nora virus infection of Drosophila melanogaster. Virus Res 175: 95–100. doi: 10.1016/j.virusres.2013.03.021
|
| [14] | Corney DC (2012) RNA-seq using next generation sequencing. Mater Method 3: 203. |
| [15] |
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25: 402–408. doi: 10.1006/meth.2001.1262
|
| [16] |
Mi H, Huang X, Muruganujan A, et al. (2017) PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res 45: D183–D189. doi: 10.1093/nar/gkw1138
|
| [17] |
Thomas PD, Kejariwal A, Guo N, et al. (2006) Applications for protein sequence-function evolution data: mRNA/protein expression analysis and coding SNP scoring tools. Nucleic Acids Res 34: W645–W650. doi: 10.1093/nar/gkl229
|
| [18] |
Huang DW, Sherman BT, Lempicki RA (2009) Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc 4: 44–57. doi: 10.1038/nprot.2008.211
|
| [19] | Huang DW, Sherman BT, Lempicki RA (2009) Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res 37: 1–13. |
| [20] |
Gramates LS, Marygold SJ, Santos GD (2017) FlyBase at 25: Looking to the future. Nucleic Acids Res 45: D663–D671. doi: 10.1093/nar/gkw1016
|
| [21] | Lemaitre Lab Uplem, List of Drosophila gene potentially involved in the immune response, 2007. Available from: https://lemaitrelab.epfl.ch/page-7767-en.html. |
| [22] | Society for Developmental Biology, the Interactive Fly: RNAi and posttranscriptional gene silencing, 2017. Available from: http://www.sdbonline.org/sites/fly/aignfam/rnaistuf.htm. |
| [23] | Smith AM, Perelson AS (2011) Influenza A virus infection kinetics: Quantitative data and models. Wires Syst Biol Med 3: 429–445. |
| [24] |
Read EL, Tovodwyer AA, Chakraborty AK (2012) Stochastic effects are important in intrahost HIV evolution even when viral loads are high. Proc Natl Acad Sci USA 109: 19727–19732. doi: 10.1073/pnas.1206940109
|
| [25] |
Pearson A, Lux A, Krieger M (1995) Expression cloning of dSR-CI, a class C macrophage-specific scavenger receptor from Drosophila melanogaster. Proc Natl Acad Sci USA 92: 4056–4060. doi: 10.1073/pnas.92.9.4056
|
| [26] |
Ligoxygakis P, Pelte N, Ji C, et al. (2002) A serpin mutant links Toll activation to melanization in the host defence of Drosophila. EMBO J 21: 6330–6337. doi: 10.1093/emboj/cdf661
|
| [27] |
Gendrin M, Zaidman-Rémy A, Broderick NA, et al. (2013) Functional analysis of PGRP-LA in Drosophila immunity. PLoS One 8: e69742. doi: 10.1371/journal.pone.0069742
|
| [28] |
Rynes J, Donohoe CD, Frommolt P, et al. (2012) Activating transcription factor 3 regulates immune and metabolic homeostasis. Mol Cell Biol 32: 3949–3962. doi: 10.1128/MCB.00429-12
|
| [29] |
Wang H, Cai Y, Chia W, et al. (2006) Drosophila homologs of mammalian TNF/TNFR-related molecules regulate segregation of Miranda/Prospero in neuroblasts. EMBO J 25: 5783–5793. doi: 10.1038/sj.emboj.7601461
|
| [30] |
Werner T, Liu G, Kang D, et al. (2000) A family of peptidoglycan recognition proteins in the fruit fly Drosophila melanogaster. Proc Natl Acad Sci USA 97: 13772–13777. doi: 10.1073/pnas.97.25.13772
|
| [31] |
Tsail CW, Mcgraw EA, Ammar ED, et al. (2008) Drosophila melanogaster mounts a unique immune response to the Rhabdovirus sigma virus. Appl Environ Microb 74: 3251–3256. doi: 10.1128/AEM.02248-07
|
| [32] |
Kim YS, Ryu JH, Han SJ, et al. (2000) Gram-negative bacteria-binding protein, a pattern recognition receptor for lipopolysaccharide and β-1,3-glucan that mediates the signaling for the induction of innate immune genes in Drosophila melanogaster cells. J Biol Chem 275: 32721–32727. doi: 10.1074/jbc.M003934200
|
| [33] |
Chamy LE, Leclerc V, Caldelari I, et al. (2008) Danger signal and PAMP sensing define binary signaling pathways upstream of Toll. Nat Immunol 9: 1165–1170. doi: 10.1038/ni.1643
|
| [34] |
Watson FL, Püttmannholgado R, Thomas F, et al. (2005) Extensive diversity of Ig-superfamily proteins in the immune system of insects. Science 309: 1874–1878. doi: 10.1126/science.1116887
|
| [35] |
Zambon RA, Nandakumar M, Vakharia VN, et al. (2005) The Toll pathway is important for an antiviral response in Drosophila. Proc Natl Acad Sci USA 102: 7257–7262. doi: 10.1073/pnas.0409181102
|
| [36] |
Grech A, Quinn R, Srinivasan D, et al. (2000) Complete structural characterisation of the mammalian and Drosophila TRAF genes: Implications for TRAF evolution and the role of RING finger splice variants. Mol Immunol 37: 721–734. doi: 10.1016/S0161-5890(00)00098-5
|
| [37] |
Cha GH, Cho KS, Lee JH, et al. (2003) Discrete functions of TRAF1 and TRAF2 in Drosophila melanogaster mediated by c-Jun N-terminal kinase and NF-κB-dependent signaling pathways. Mol Cell Biol 23: 7982–7991. doi: 10.1128/MCB.23.22.7982-7991.2003
|
| [38] |
Berkey CD, Blow N, Watnick PI (2009) Genetic analysis of Drosophila melanogaster susceptibility to intestinal Vibrio cholerae infection. Cell Microbiol 11: 461–474. doi: 10.1111/j.1462-5822.2008.01267.x
|
| [39] |
Moy RH, Gold B, Molleston JM, et al. (2014) Antiviral autophagy restricts Rift Valley fever virus infection and is conserved from flies to mammals. Immunity 40: 51–65. doi: 10.1016/j.immuni.2013.10.020
|
| [40] |
Xu YC, Wu RF, Gu Y, et al. (2002) Involvement of TRAF4 in oxidative activation of c-Jun N-terminal kinase. J Biol Chem 277: 28051–28057. doi: 10.1074/jbc.M202665200
|
| [41] |
Ha EM, Oh CT, Bae YS, et al. (2005) A direct role for dual oxidase in Drosophila gut immunity. Science 310: 847–850. doi: 10.1126/science.1117311
|
| [42] |
Tian C, Gao B, Rodriguez MC, et al. (2008) Gene expression, antiparasitic activity, and functional evolution of the drosomycin family. Mol Immunol 45: 3909–3916. doi: 10.1016/j.molimm.2008.06.025
|
| [43] |
Lematire B, Nicolas E, Michaut L, et al. (1996) The dorsoventral regulatory gene cassette spätzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 86: 973–983. doi: 10.1016/S0092-8674(00)80172-5
|
| [44] |
Kurucz E, Markus R, Zsamboki J, et al. (2007) Nimrod, a putative phagocytosis receptor with EGF repeats in Drosophila plasmatocytes. Curr Biol 17: 649–654. doi: 10.1016/j.cub.2007.02.041
|
| [45] |
Van Mierlo JT, Bronkhorst AW, Overheul GJ, et al. (2012) Convergent evolution of Argonaute-2 slicer antagonism in two distinct insect RNA viruses. PLoS Pathog 8: e1002872. doi: 10.1371/journal.ppat.1002872
|
| [46] |
Kemp C, Imler JL (2009) Antiviral immunity in Drosophila. Curr Opin Immunol 21: 3–9. doi: 10.1016/j.coi.2009.01.007
|
| [47] |
Baker DA, Russell S (2009) Gene expression during Drosophila melanogaster egg development before and after reproductive diapause. BMC Genomics 10: 1–16. doi: 10.1186/1471-2164-10-1
|
| [48] | Potempa J, Korzus E, Travis J (1994) The serpin superfamily of proteinase inhibitors: Structure, function, and regulation. J Biol Chem 269: 15957–15960. |
| [49] |
Irving P, Trozler L, Heuer TS, et al. (2001) A genome-wide analysis of immune responses in Drosophila. Proc Natl Acad Sci USA 98: 15119–15124. doi: 10.1073/pnas.261573998
|
| [50] |
Tanj T, Yun EY, Ip YT (2010) Heterodimers of NF-κB transcription factors DIF and Relish regulate antimicrobial peptide genes in Drosophila. Proc Natl Acad Sci USA 107: 14715–14720. doi: 10.1073/pnas.1009473107
|
 microbiol-04-01-123-s01.pdf microbiol-04-01-123-s01.pdf |
 |
| microbiol-04-01-123-s02.xlsx |
|
| microbiol-04-01-123-s03.xlsx |
|
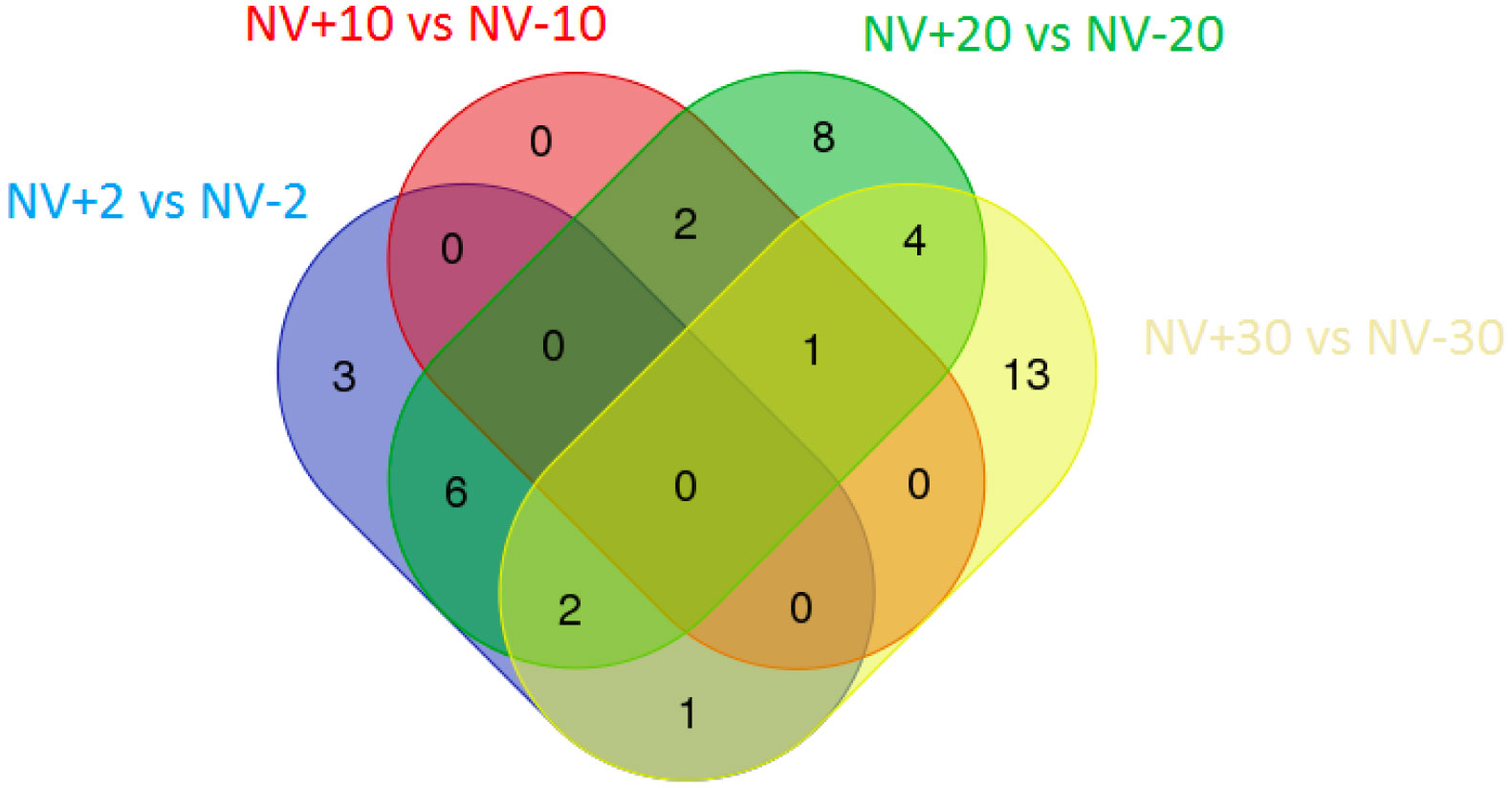
Figures(1) / Tables(6)

Wilfredo Lopez, Alexis M. Page, Darby J. Carlson, Brad L. Ericson, Matyas F. Cserhati, Chittibabu Guda, Kimberly A. Carlson. Analysis of immune-related genes during Nora virus infection of Drosophila melanogaster using next generation sequencing[J]. AIMS Microbiology, 2018, 4(1): 123-139. doi: 10.3934/microbiol.2018.1.123







 DownLoad:
DownLoad: