Citation: Katie Lewis, Jamie Foster, Frank Hons, Thomas Boutton. Initial aggregate formation and soil carbon storage from lipid-extracted algae amendment[J]. AIMS Environmental Science, 2017, 4(6): 743-762. doi: 10.3934/environsci.2017.6.743

| [1] | Schlesinger WH (1997) Biogeochemistry. Geotimes 42: 44. |
| [2] |
Amundson R (2001) The carbon budget in soils. Annu Rev Earth Planet Sci 29: 535-562. doi: 10.1146/annurev.earth.29.1.535

|
| [3] |
Degens BP, Schipper LA, Sparling GP, et al. (2000) Decreases in organic C reserves in soils can reduce the catabolic diversity of soil microbial communities. Soil Biol Biochem 32: 189-196. doi: 10.1016/S0038-0717(99)00141-8
|
| [4] |
Karami A, Homaee M, Afzalinia S, et al. (2012) Organic resource management: impacts on soil aggregate stability and other soil physico-chemical properties. Agric Ecosyst Environ 148: 22-28. doi: 10.1016/j.agee.2011.10.021
|
| [5] |
Plante AF, McGill WB (2002) Soil aggregate dynamics and the retention of organic matter in laboratory-incubated soil with differing simulated tillage frequencies. Soil Till Res 66: 79-92. doi: 10.1016/S0167-1987(02)00015-6
|
| [6] |
Six J, Conant RT, Paul EA, et al. (2002) Stabilization mechanisms of soil organic matter: implication for C-saturation of soils. Plant Soil 241: 155-176. doi: 10.1023/A:1016125726789
|
| [7] |
Tisdall JM, Oades JM (1982) Organic matter and water-stable aggregates in soils. J Soil Sci 33: 141-163. doi: 10.1111/j.1365-2389.1982.tb01755.x
|
| [8] |
Chivenge P, Vanlauwe B, Gentile R, et al. (2011) Comparison of organic versus mineral resource effects on short-term aggregate carbon and nitrogen dynamics in a sandy soil versus a fine textured soil. Agric Ecosyst Environ 140: 361-371. doi: 10.1016/j.agee.2010.12.004
|
| [9] |
Six J, Elliott ET, Paustian K (1999) Aggregate and soil organic matter dynamics under conventional and no-tillage systems. Soil Sci Soc Am J 63: 1350-1358. doi: 10.2136/sssaj1999.6351350x
|
| [10] |
Jastrow JD, Boutton TW, Miller RM (1996) Carbon dynamics of aggregate-associated organic matter estimated by carbon-13 natural abundance. Soil Sci Soc Am J 60: 801-807. doi: 10.2136/sssaj1996.03615995006000030017x
|
| [11] | Wright AL, Hons FM (2005) Soil carbon and nitrogen storage in aggregates from different tillage and crop regimes. Soil Sci Soc Am J 69: 141-147. |
| [12] |
Ghidey F, Alberts EE (1993) Residue type and placement effects on decomposition: field study and model evaluation. Trans ASAE 36: 1611-1617. doi: 10.13031/2013.28502
|
| [13] |
Franzluebbers AJ, Zuberer DA, Hons FM (1995) Comparison of microbiological methods for evaluating quality and fertility of soil. Biol Fert Soils 19: 135-140. doi: 10.1007/BF00336149
|
| [14] | Rothlisberger-Lewis KL, Foster JL, Hons FM (2015) Soil carbon and nitrogen dynamics as affected by lipid-extracted algae application. Geoderma 262: 140-146. |
| [15] |
Gelin F, Volkman J, Largeau C, et al. (1999) Distribution of aliphatic, nonhydrolyzable biopolymers in marine microalgae. Org Geochem 30: 147-159. doi: 10.1016/S0146-6380(98)00206-X
|
| [16] |
Poirier N, Derenne S, Rouzaud J, et al. (2000) Chemical structure and sources of the macromolecular, resistant, organic fraction isolated from a forest soil (Lacadee, south-west France). Org Geochem 31: 813-827. doi: 10.1016/S0146-6380(00)00067-X
|
| [17] |
Balesdent J, Mariotti A (1987) Natural 13C abundance as a tracer for studies of soil organic matter dynamics. Soil Biol Biochem 19: 25-30. doi: 10.1016/0038-0717(87)90120-9
|
| [18] | Tieszen TJ, Boutton TW (1989) Stable carbon isotopes in terrestrial ecosystem research, In: Rundel PW et al. Stable isotopes in ecological research, Ecological studies 68, New York: Springer-Verlag, 167-195. |
| [19] | Boutton TW (1996) Stable carbon isotope ratios of soil organic matter and their use as indicators of vegetation and climate change, In: Boutton TW, Yamakaki S, Mass spectrometry of soils, New York: Marcel Dekker, 47-82. |
| [20] |
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol Plant Molec Biol 40: 503-537. doi: 10.1146/annurev.pp.40.060189.002443
|
| [21] | Wada E, Ando T, Kumazawa K (1995) Biodiversity of stable isotope ratios, In: Wada E et al., Stable isotope in the biosphere, Japan: Kyoto University Press, 7-14. |
| [22] |
Gerzabek MH, Pichlmayer F, Kirchmann H, et al. (1997) The response of soil organic matter to manure amendments in a long-term experiment in Ultuna, Sweden. Euro J Soil Sci 48: 273-282. doi: 10.1111/j.1365-2389.1997.tb00547.x
|
| [23] |
O'Leary MH (1981) Carbon isotope fractionation in plants. Phytochem 20: 553-567. doi: 10.1016/0031-9422(81)85134-5
|
| [24] |
Solomon D, Fritzche F, Lehmann J, et al. (2002) Soil organic matter dynamics in the subhumid agroecosystems of the Ethiopian highlands: evidence from natural 13C abundance and particle-size fractionation. Soil Sci Soc Am J 66: 969-978. doi: 10.2136/sssaj2002.9690
|
| [25] |
Balesdent J, Mariotti A (1987) Natural 13C abundance as a tracer for studies of soil organic matter dynamics. Soil Biol Biochem 19: 25-30. doi: 10.1016/0038-0717(87)90120-9
|
| [26] |
Liao JD, Boutton TW, Jastrow JD (2006) Organic matter turnover in soil physical fractions following woody plant invasion of grassland: evidence from natural 13C and 15N. Soil Biol Biochem 38: 3197-3210. doi: 10.1016/j.soilbio.2006.04.004
|
| [27] |
Storer DA (1984) A simple high volume ashing procedure for determining soil organic matter. Comm Soil Sci Plant Anal 15: 759-772. doi: 10.1080/00103628409367515
|
| [28] | McGeehan SL, Naylor DV (1998) Automated instrumental analysis of carbon and nitrogen in plant and soil samples. Comm Soil Sci Plant Anal 19: 493. |
| [29] | Schulte EE, Hopkins BG (1996) Estimation of soil organic matter by weight by weight loss-on-ignition, In: Magdoff FR, Tabatabai MA, Hanlon EA, Jr, Soil organic matter: analysis and interpretation, Wisconsin: Soil Sci Soc Am, 21-32. |
| [30] | Isaac RA, Johnson WC (1975) Collaborative study of wet and dry ashing techniques for the elemental analysis of plant tissue by atomic absorption spectrophotometry. J Assoc Off Anal Chem 58: 436-440. |
| [31] | Havlin JL, Soltanpour PN (1989) A nitric acid and plant digest method for use with inductively coupled plasma spectrometry. Commun Soil Sci Plant Anal 14: 969-980. |
| [32] | Van Soest PJ, Robertson JB, Lewis BA (1991) Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J Dairy Sci 74: 35-68. |
| [33] | AOAC (1990) Official methods of analysis. 15th ed, Virginia: Assoc Off Anal Chem. |
| [34] | Ankom, 2013. Method 8-determining acid detergent lignin in beakers. Available from: http://www.ankom.com/media/documents/Method_8_Lignin_in_beakers_3_13_13.pdf. |
| [35] |
Mehlich A (1978) New extractant for soil test evaluation of phosphorus, potassium, magnesium, calcium, sodium, manganese, and zinc. Comm Soil Sci Plant Anal 9: 477-492. doi: 10.1080/00103627809366824
|
| [36] |
Mehlich A (1984) Mehlich-III soil test extractant: a modification of Mehlich-II extractant. Comm Soil Sci Plant Anal 15: 1409-1416. doi: 10.1080/00103628409367568
|
| [37] |
Lindsay WL, Norvell WA (1978) Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci Soc Am J 42: 421-428. doi: 10.2136/sssaj1978.03615995004200030009x
|
| [38] |
Chepil WS, Bisal F (1943) A rotary sieve method for determining the size distribution of soil clods. Soil Sci 56: 95-100. doi: 10.1097/00010694-194308000-00002
|
| [39] | Kemper WD, Chepil WS (1965) Size distribution of aggregates, In: Black CA, et al., Methods of soil analysis, part I. 9th ed. Wisconsin: Am Soc Agron, 499-510. |
| [40] | Rhoades JD (1982) Soluble salts, In: Page AL, et al., Methods of soil analysis: part 2. Agronomy Monogr 9. 2nd ed. Wisconsin: Am Soc Agron and Soil Sci Soc Am, 167-178. |
| [41] |
Coplen TB (1996) New guidelines for reporting stable hydrogen, carbon, and oxygen isotope-ratio data. Geochim Cosmochim Acta 60: 3359-3360. doi: 10.1016/0016-7037(96)00263-3
|
| [42] | SAS Institute (2012) SAS system for Windows. Release 9.3, SAS Inst Inc, Cary, North Carolina USA. |
| [43] |
Chivenge P, Vanlauwe B, Gentile R, et al. (2011). Organic resource quality influences short-term aggregate dynamics and soil organic carbon and nitrogen accumulation. Soil Biol Biochem 43: 657-666. doi: 10.1016/j.soilbio.2010.12.002
|
| [44] |
Whalen JK, Chang C (2002) Macroaggregate characteristics in cultivated soils after 25 annual manure applications. Soil Sci Soc Am J 66: 1637-1647. doi: 10.2136/sssaj2002.1637
|
| [45] | Khaleel R, Reddy KR, Overcash MR (1981) Changes in soil physical properties due to organic waste applications: a review. J Environ Qual 10: 133-141. |
| [46] |
Sun H, Larney FJ, Bullock MS (1995) Soil amendments and water-stable aggregation of a desurfaced dark brown chernozem. Can J Soil Sci 75: 319-325. doi: 10.4141/cjss95-046
|
| [47] |
Lynch DH, Voroney RP, Warman PR (2006) Use of 13C and 15N natural abundance techniques to characterize carbon and nitrogen dynamics in composting and in compost-amended soils. Soil Biol Biochem 38: 103-114. doi: 10.1016/j.soilbio.2005.04.022
|
| [48] |
Zhang H, Ding W, Luo J, et al. (2015) The dynamics of glucose-derived 13C incorporation into aggregates of a sandy loam soil following two-decade compost or inorganic fertilizer amendments. Soil Till Res 148: 14-19. doi: 10.1016/j.still.2014.11.010
|
| [49] |
Werth M, Kuzyakov Y (2010) 13C fractionation at the root-microorganisms-soil interface: A review and outlook for partitioning studies. Soil Biol Biochem 42: 1372-1384. doi: 10.1016/j.soilbio.2010.04.009
|
| [50] |
Cambardella CA, Elliott ET (1993). Carbon and nitrogen distribution in aggregates from cultivated and native grassland soils. Soil Sci Soc Am J 57: 1071-1076. doi: 10.2136/sssaj1993.03615995005700040032x
|
| [51] |
Bhattacharyya R, Prakash V, Kundu S, et al. (2010) Long-term effects of fertilization on carbon and nitrogen sequestration and aggregate associated carbon and nitrogen in the Indian sub-Himalayas. Nutr Cycl Agroecosyst 86: 1-16. doi: 10.1007/s10705-009-9270-y
|
| [52] |
Udom BE, Nuga BO, Adesodun JK (2016) Water-stable aggregates and aggregate-associated organic carbon and nitrogen after three annual applications of poultry manure and spent mushroom wastes. Appl Soil Ecol 101: 5-10. doi: 10.1016/j.apsoil.2016.01.007
|
| [53] |
Yousaf B, Liu G, Wang R, et al. (2017) Investigating the biochar effects on C-mineralization and sequestration of carbon in soil compared with conventional amendments using the stable isotope (delta C-13) approach. Glob Change Biol Bioenergy 9: 1085-1099. doi: 10.1111/gcbb.12401
|
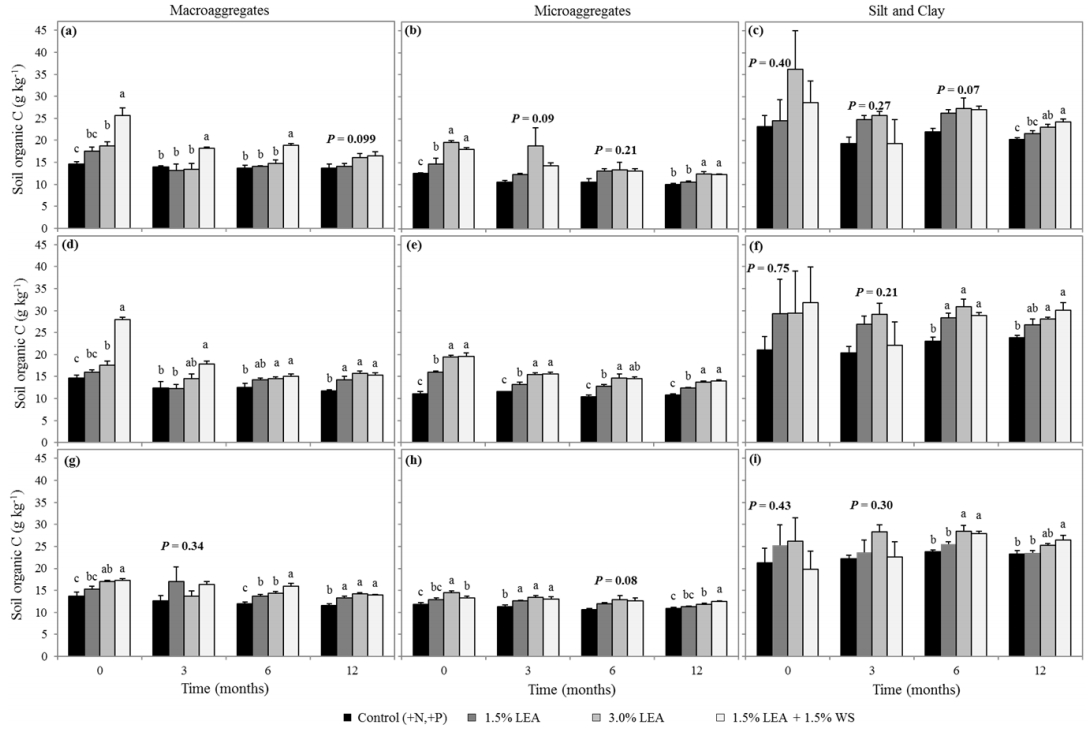
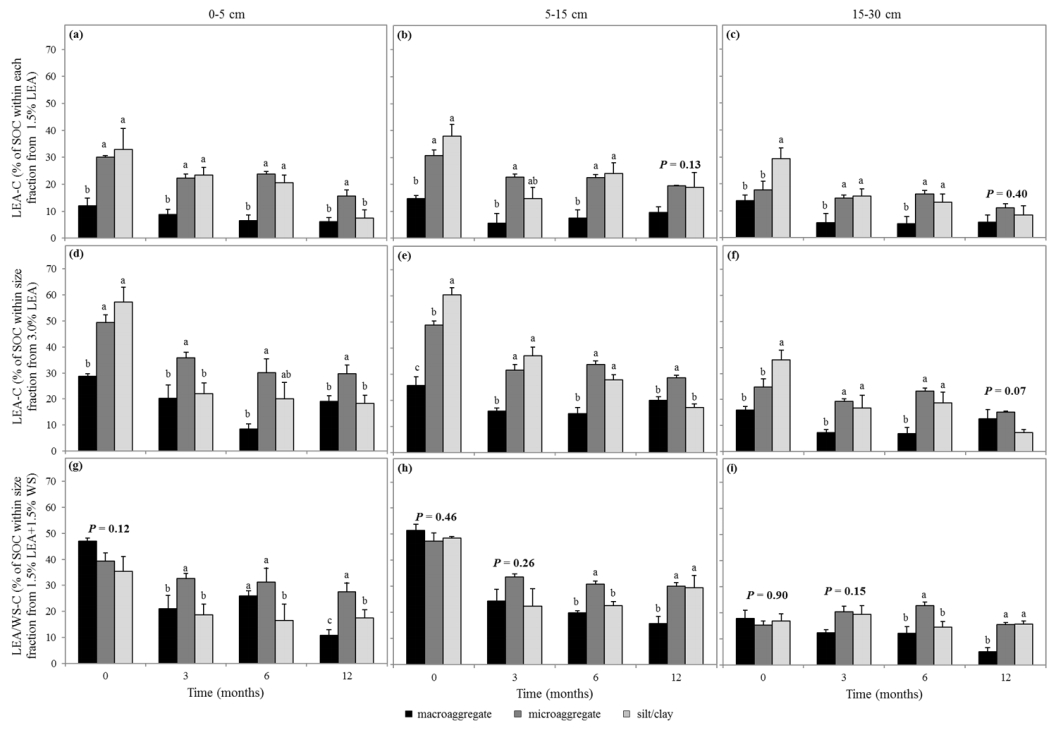
Figures(6) / Tables(3)

Katie Lewis, Jamie Foster, Frank Hons, Thomas Boutton. Initial aggregate formation and soil carbon storage from lipid-extracted algae amendment[J]. AIMS Environmental Science, 2017, 4(6): 743-762. doi: 10.3934/environsci.2017.6.743







 DownLoad:
DownLoad: