Citation: Katie M. Dunleavy, Eugene Milshteyn, Zachary Sorrentino, Natasha L. Pirman, Zhanglong Liu, Matthew B. Chandler, Peter W. D’Amore, Gail E. Fanucci. Spin-label scanning reveals conformational sensitivity of the bound helical interfaces of IA3[J]. AIMS Biophysics, 2018, 5(3): 166-181. doi: 10.3934/biophy.2018.3.166

| [1] |
Dunker AK, Lawson JD, Brown CJ, et al. (2001) Intrinsically disordered protein. J Mol Graph Model 19: 26–59. doi: 10.1016/S1093-3263(00)00138-8

|
| [2] |
Tompa P (2002) Intrinsically unstructured proteins. Trends Biochem Sci 27: 527–533. doi: 10.1016/S0968-0004(02)02169-2
|
| [3] |
Wright PE, Dyson HJ (1999) Intrinsically unstructured proteins: re-assessing the protein structure-function paradigm. J Mol Biol 293: 321–331. doi: 10.1006/jmbi.1999.3110
|
| [4] |
Dyson HJ, Wright PE (2005) Intrinsically unstructured proteins and their functions. Nat Rev Mol Cell Biol 6: 197–208. doi: 10.1038/nrm1589
|
| [5] |
Morin B, Bourhis JM, Belle V, et al. (2006) Assessing induced folding of an intrinsically disordered protein by site-directed spin-labeling electron paramagnetic resonance spectroscopy. J Phys Chem B 110: 20596–20608. doi: 10.1021/jp063708u
|
| [6] |
Knott M, Best RB (2012) A preformed binding interface in the unbound ensemble of an intrinsically disordered protein: evidence from molecular simulations. PLoS Comput Biol 8: e1002605. doi: 10.1371/journal.pcbi.1002605
|
| [7] | Dunker AK, Oldfield CJ, Meng J, et al. (2008) The unfoldomics decade: an update on intrinsically disordered proteins. BMC Genomics 9: S1. |
| [8] |
Dyson HJ, Wright PE (2002) Coupling of folding and binding for unstructured proteins. Curr Opin Struct Biol 12: 54–60. doi: 10.1016/S0959-440X(02)00289-0
|
| [9] |
Fuxreiter M, Simon I, Friedrich P, et al. (2004) Preformed structural elements feature in partner recognition by intrinsically unstructured proteins. J Mol Biol 338: 1015–1026. doi: 10.1016/j.jmb.2004.03.017
|
| [10] |
Pancsa R, Tompa P (2012) Structural disorder in eukaryotes. PLoS One 7: e34687. doi: 10.1371/journal.pone.0034687
|
| [11] |
Li M, Phylip LH, Lees WE, et al. (2000) The aspartic proteinase from Saccharomyces cerevisiae folds its own inhibitor into a helix. Nat Struct Biol 7: 113–117. doi: 10.1038/72378
|
| [12] |
Phylip LH, Lees WE, Brownsey BG, et al. (2001) The potency and specificity of the interaction between the IA3 inhibitor and its target aspartic proteinase from Saccharomyces cerevisiae. J Biol Chem 276: 2023–2030. doi: 10.1074/jbc.M008520200
|
| [13] |
Green TB, Ganesh O, Perry K, et al. (2004) IA3, an aspartic proteinase inhibitor from Saccharomyces cerevisiae, is intrinsically unstructured in solution. Biochemistry 43: 4071–4081. doi: 10.1021/bi034823n
|
| [14] |
Ganesh OK, Green TB, Edison AS, et al. (2006) Characterizing the residue level folding of the intrinsically unstructured IA3. Biochemistry 45: 13585–13596. doi: 10.1021/bi061358w
|
| [15] |
Narayanan R, Ganesh OK, Edison AS, et al. (2008) Kinetics of folding and binding of an intrinsically disordered protein: the inhibitor of yeast aspartic proteinase YPrA. J Am Chem Soc 130: 11477–11485. doi: 10.1021/ja803221c
|
| [16] |
Wang J, Wang Y, Chu X, et al. (2011) Multi-scaled explorations of binding-induced folding of intrinsically disordered protein inhibitor IA3 to its target enzyme. PLoS Comput Biol 7: e1001118. doi: 10.1371/journal.pcbi.1001118
|
| [17] |
Pirman NL, Milshteyn E, Galiano L, et al. (2011) Characterization of the disordered-to-alpha-helical transition of IA(3) by SDSL-EPR spectroscopy. Protein Sci 20: 150–159. doi: 10.1002/pro.547
|
| [18] |
Le Breton N, Martinho M, Kabytaev K, et al. (2014) Diversification of EPR signatures in Site Directed Spin Labeling using a beta-phosphorylated nitroxide. Phys Chem Chem Phys 16: 4202–4209. doi: 10.1039/c3cp54816c
|
| [19] |
Hubbell WL, Cafiso DS, Altenbach C (2000) Identifying conformational changes with site-directed spin labeling. Nat Struct Biol 7: 735–739. doi: 10.1038/78956
|
| [20] |
Hubbell WL, McHaourab HS, Altenbach C, et al. (1996) Watching proteins move using site-directed spin labeling. Structure 4: 779–783. doi: 10.1016/S0969-2126(96)00085-8
|
| [21] |
Fanucci GE, Cafiso DS (2006) Recent advances and applications of site-directed spin labeling. Curr Opin Struct Biol 16: 644–653. doi: 10.1016/j.sbi.2006.08.008
|
| [22] |
Longhi S, Belle V, Fournel A, et al. (2011) Probing structural transitions in both structured and disordered proteins using site-directed spin-labeling EPR spectroscopy. J Pept Sci 17: 315–328. doi: 10.1002/psc.1344
|
| [23] |
Cafiso DS (2014) Identifying and quantitating conformational exchange in membrane proteins using site-directed spin labeling. Accounts Chem Res 47: 3102–3109. doi: 10.1021/ar500228s
|
| [24] |
Fleissner MR, Brustad EM, Kalai T, et al. (2009) Site-directed spin labeling of a genetically encoded unnatural amino acid. P Natl Acad Sci USA 106: 21637–21642. doi: 10.1073/pnas.0912009106
|
| [25] |
Lorenzi M, Puppo C, Lebrun R, et al. (2011) Tyrosine-targeted spin labeling and EPR spectroscopy: an alternative strategy for studying structural transitions in proteins. Angew Chem Int Edit 50: 9108–9111. doi: 10.1002/anie.201102539
|
| [26] |
Mileo E, Etienne E, Martinho M, et al. (2013) Enlarging the panoply of site-directed spin labeling electron paramagnetic resonance (SDSL-EPR): sensitive and selective spin-labeling of tyrosine using an isoindoline-based nitroxide. Bioconjugate Chem 24: 1110–1117. doi: 10.1021/bc4000542
|
| [27] |
Mileo E, Lorenzi M, Erales J, et al. (2013) Dynamics of the intrinsically disordered protein CP12 in its association with GAPDH in the green alga Chlamydomonas reinhardtii: a fuzzy complex. Mol Biosyst 9: 2869–2876. doi: 10.1039/c3mb70190e
|
| [28] |
Columbus L, Hubbell WL (2002) A new spin on protein dynamics. Trends Biochem Sci 27: 288–295. doi: 10.1016/S0968-0004(02)02095-9
|
| [29] | McHaourab HS, Lietzow MA, Hideg K, et al. (1996) Motion of spin-labeled side chains in T4 lysozyme. Correlation with protein structure and dynamics. Biochemistry 35: 7692–7704. |
| [30] |
Qin PZ, Iseri J, Oki A (2006) A model system for investigating lineshape/structure correlations in RNA site-directed spin labeling. Biochem Bioph Res Co 343: 117–124. doi: 10.1016/j.bbrc.2006.02.138
|
| [31] |
Bartelli NL, Hazelbauer GL (2011) Direct evidence that the carboxyl-terminal sequence of a bacterial chemoreceptor is an unstructured linker and enzyme tether. Protein Sci 20: 1856–1866. doi: 10.1002/pro.719
|
| [32] |
Belle V, Rouger S, Costanzo S, et al. (2008) Mapping alpha-helical induced folding within the intrinsically disordered C-terminal domain of the measles virus nucleoprotein by site-directed spin-labeling EPR spectroscopy. Proteins 73: 973–988. doi: 10.1002/prot.22125
|
| [33] |
Frazier AA, Wisner MA, Malmberg NJ, et al. (2002) Membrane orientation and position of the C2 domain from cPLA2 by site-directed spin labeling. Biochemistry 41: 6282–6292. doi: 10.1021/bi0160821
|
| [34] |
Fanucci GE, Coggshall KA, Cadieux N, et al. (2003) Substrate-induced conformational changes of the periplasmic N-terminus of an outer-membrane transporter by site-directed spin labeling. Biochemistry 42: 1391–1400. doi: 10.1021/bi027120z
|
| [35] |
Kim M, Fanucci GE, Cafiso DS (2007) Substrate-dependent transmembrane signaling in TonB-dependent transporters is not conserved. P Natl Acad Sci USA 104: 11975–11980. doi: 10.1073/pnas.0702172104
|
| [36] |
Esquiaqui JM, Sherman EM, Ye JD, et al. (2016) Conformational flexibility and dynamics of the internal loop and helical regions of the kink-Turn Motif in the glycine riboswitch by site-directed spin-labeling. Biochemistry 55: 4295–4305. doi: 10.1021/acs.biochem.6b00287
|
| [37] |
Hirst JD, Brooks III CL (1994) Helicity, circular dichroism and molecular dynamics of proteins. J Mol Biol 243: 173–178. doi: 10.1006/jmbi.1994.1644
|
| [38] | Greenfield NJ (2006) Using circular dichroism spectra to estimate protein secondary structure. Nat Protoc 1: 2876–2890. |
| [39] |
Columbus L, Hubbell WL (2004) Mapping backbone dynamics in solution with site-directed spin labeling: GCN4-58 bZip free and bound to DNA. Biochemistry 43: 7273–7287. doi: 10.1021/bi0497906
|
 biophy-05-03-166-s001.pdf biophy-05-03-166-s001.pdf |
 |
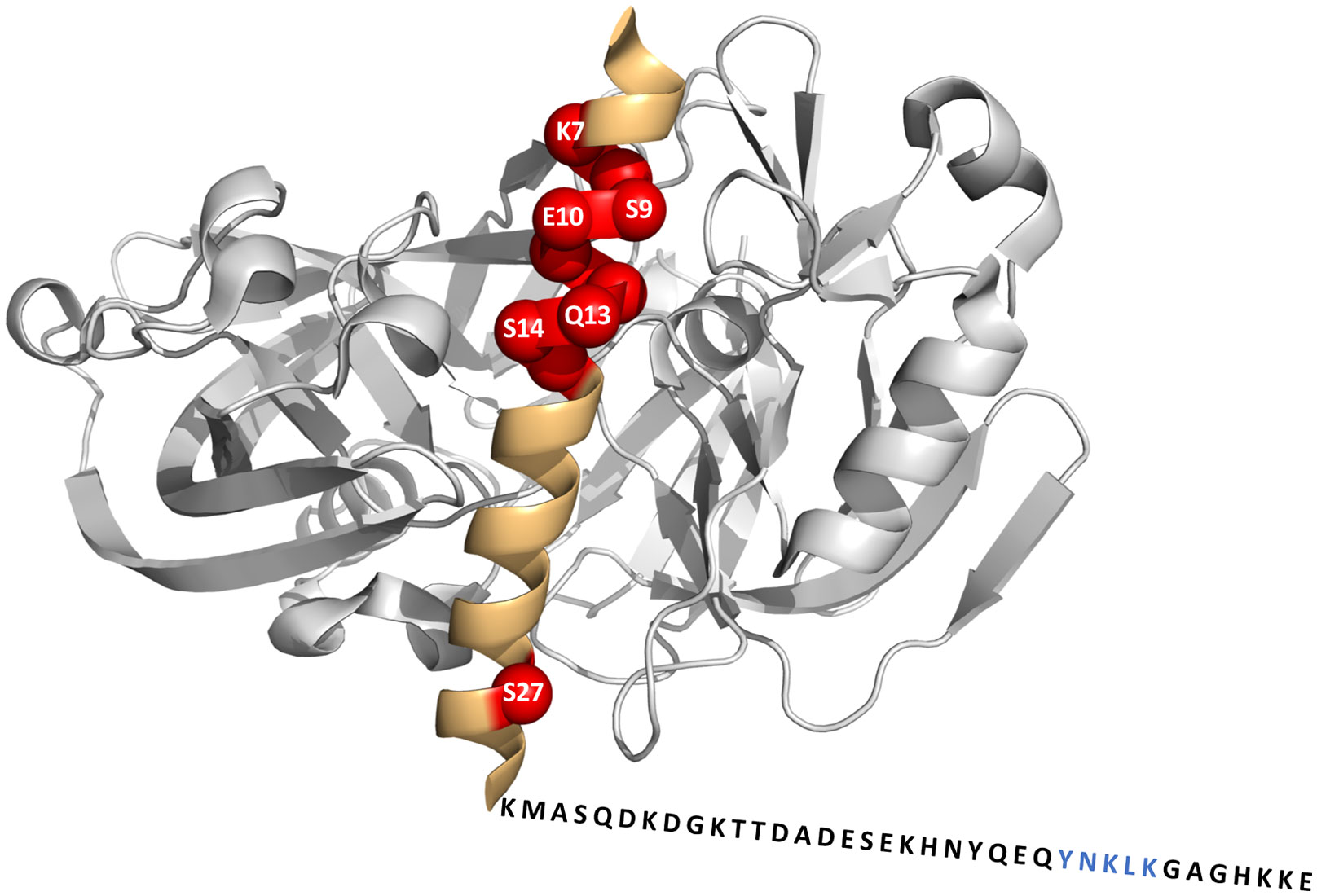
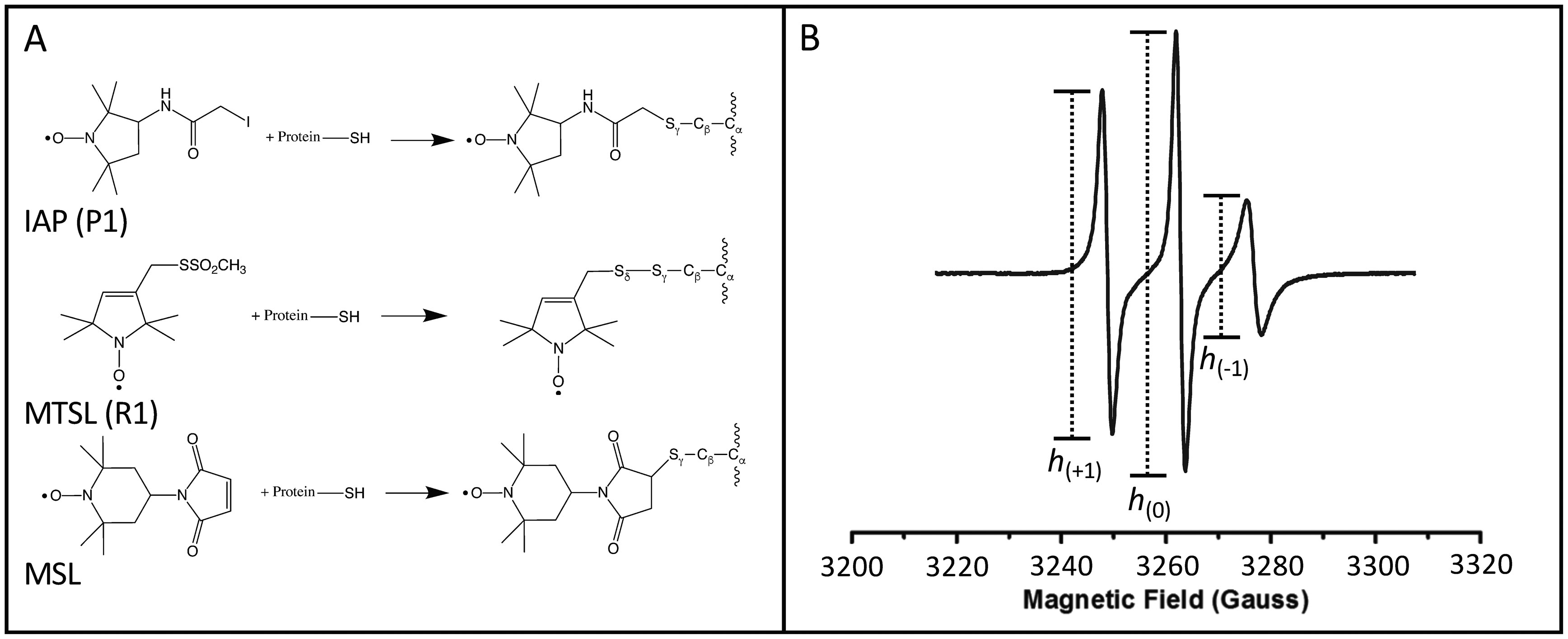
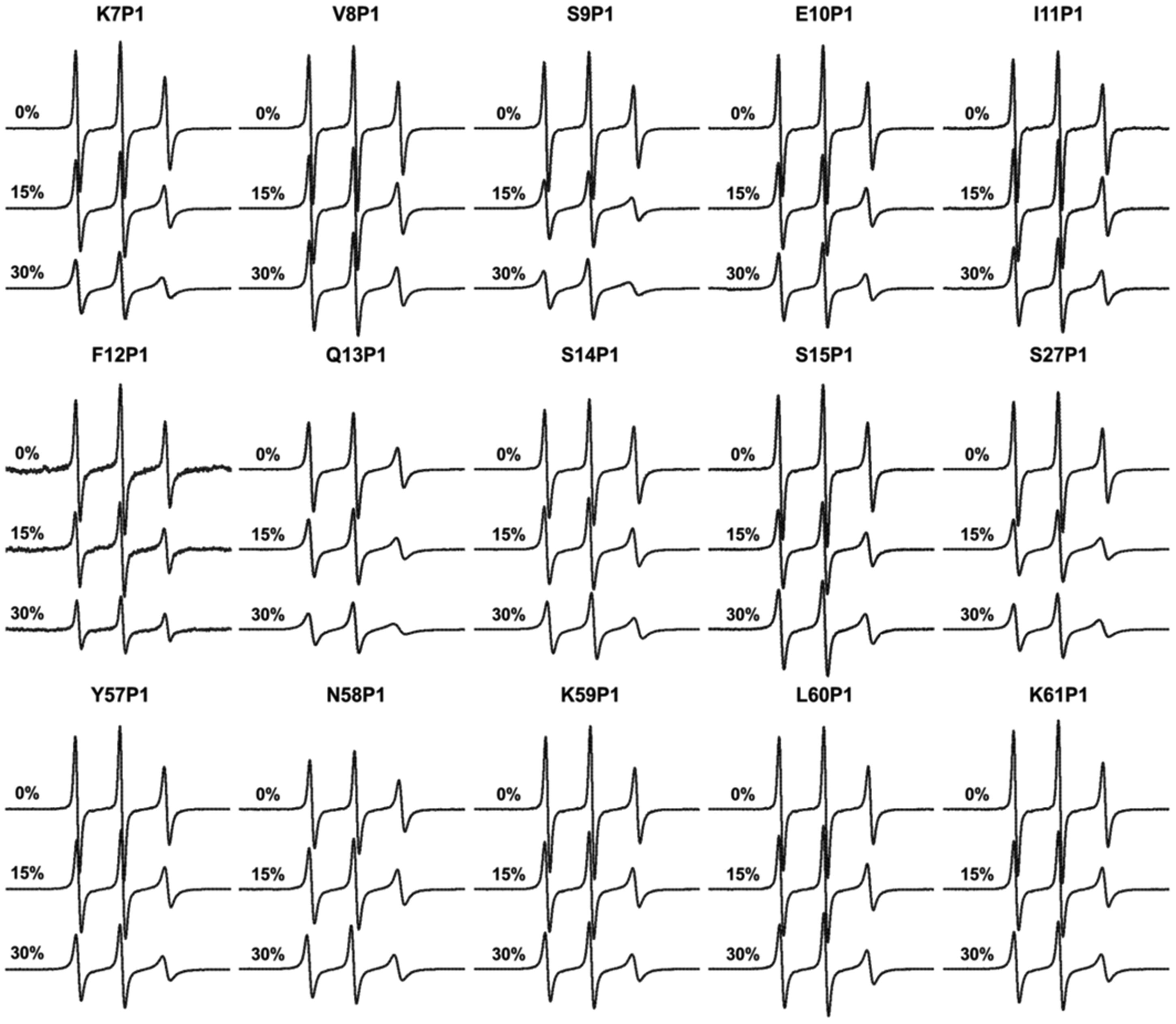
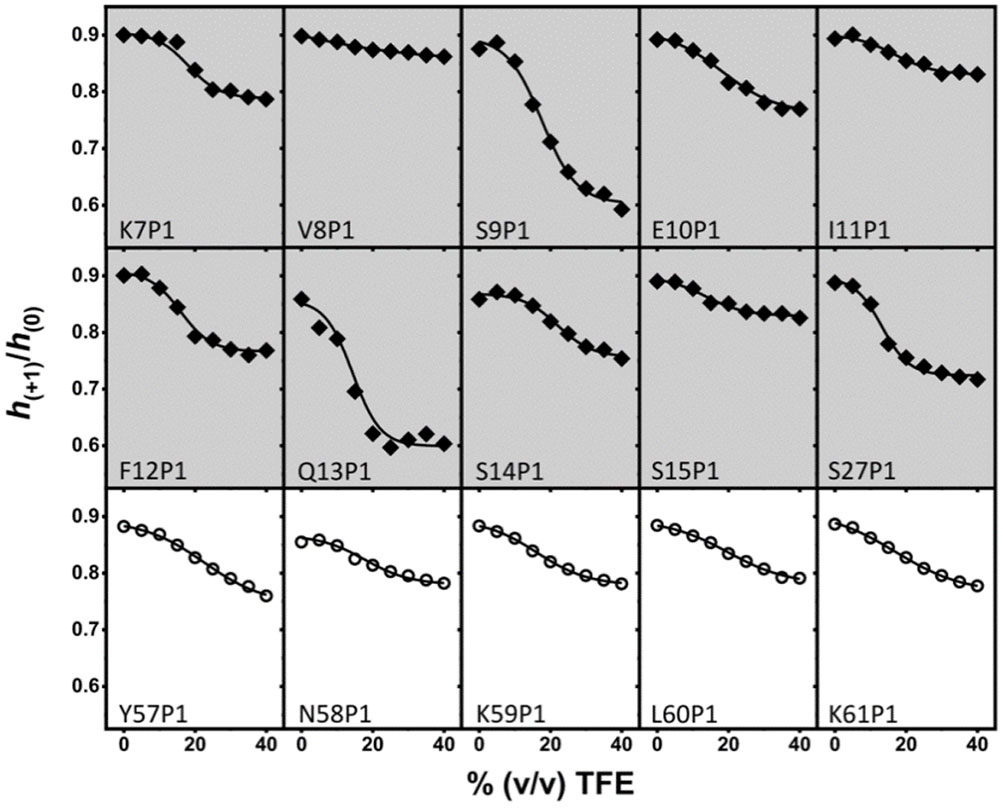
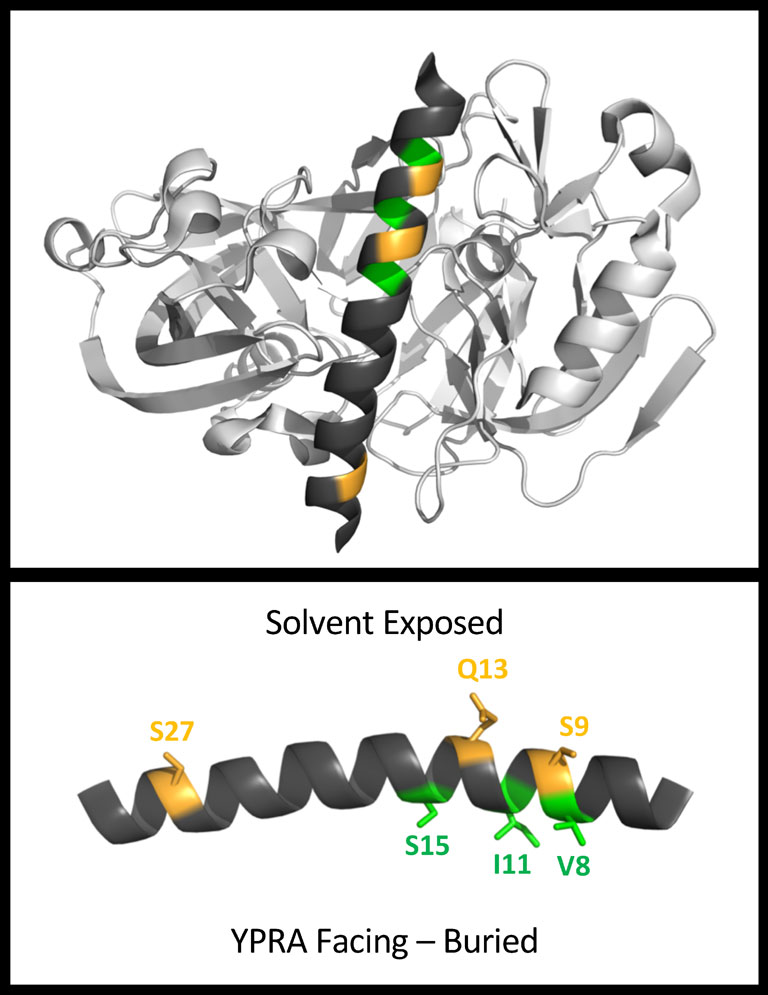
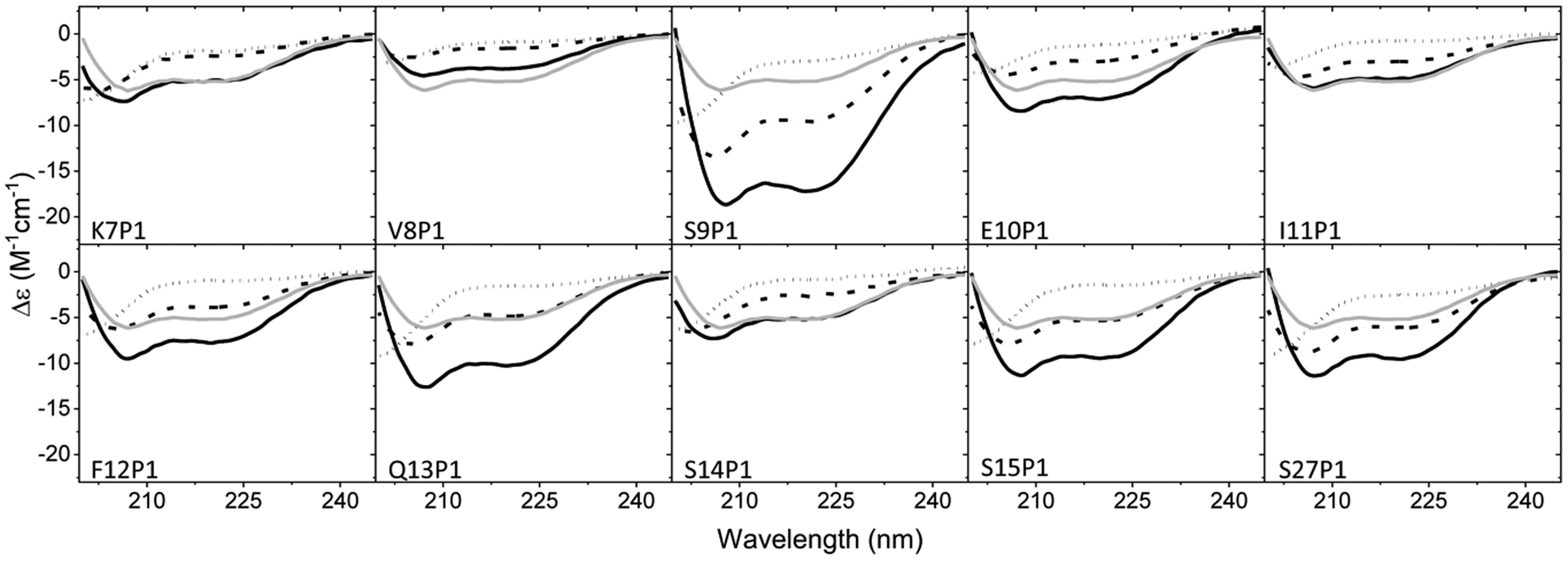
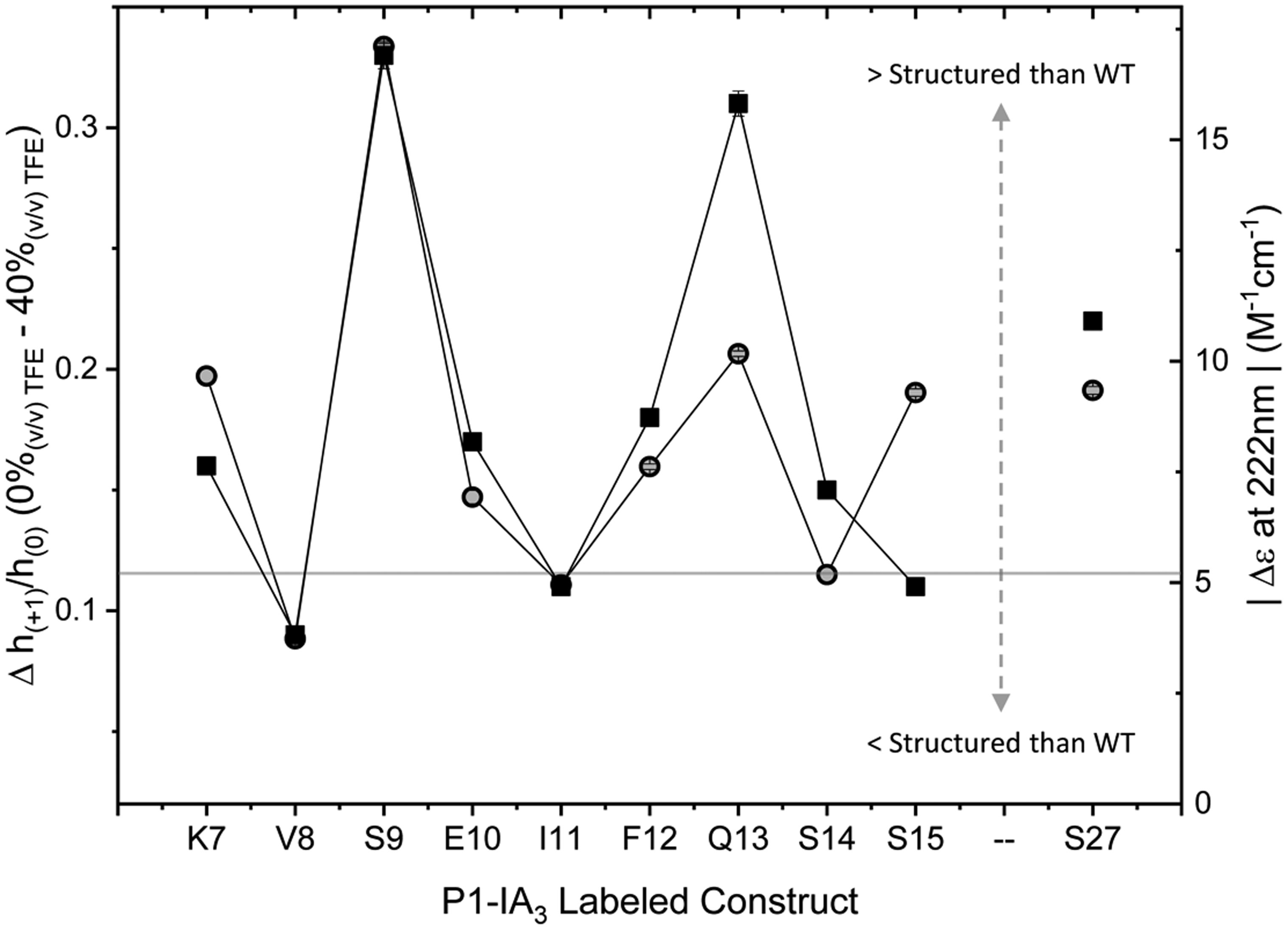
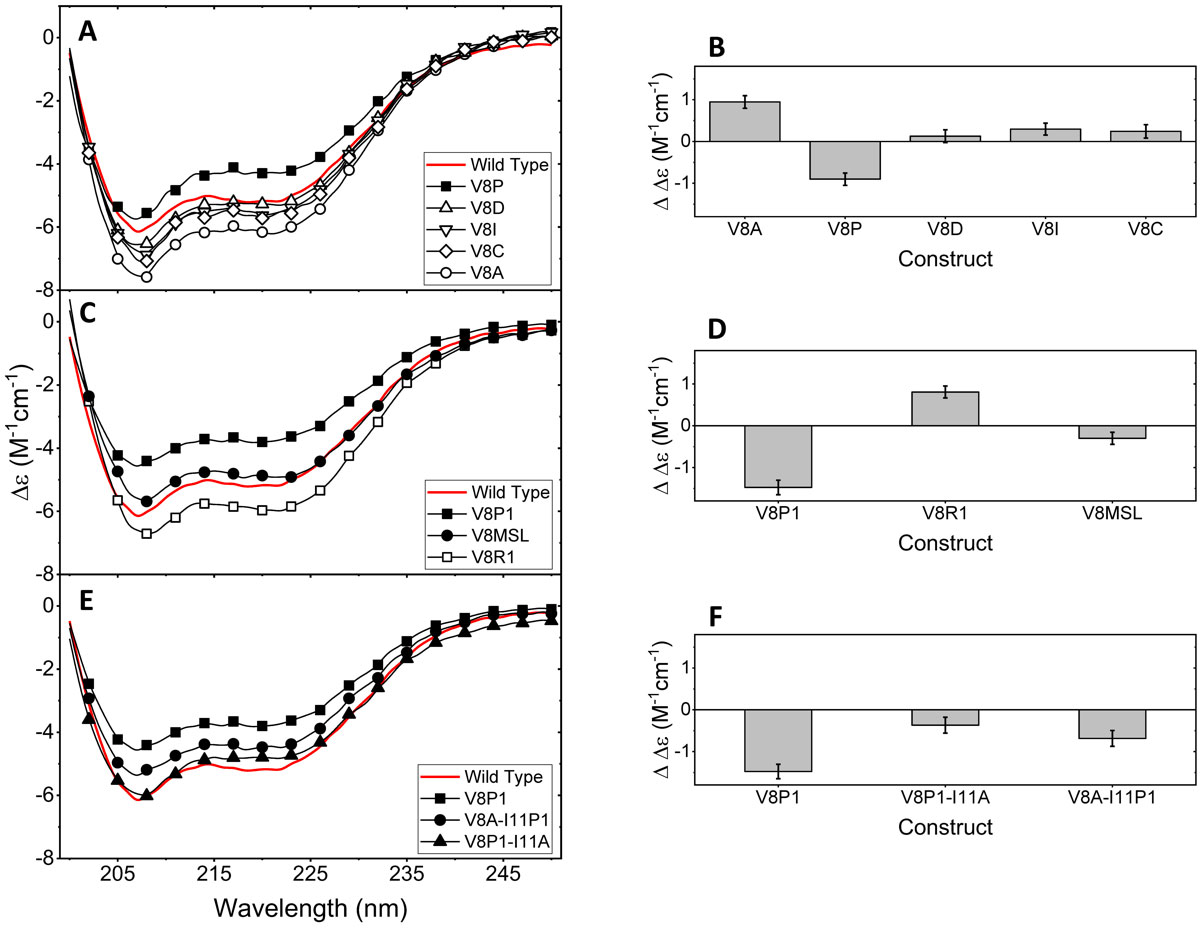
Figures(8)

Katie M. Dunleavy, Eugene Milshteyn, Zachary Sorrentino, Natasha L. Pirman, Zhanglong Liu, Matthew B. Chandler, Peter W. D’Amore, Gail E. Fanucci. Spin-label scanning reveals conformational sensitivity of the bound helical interfaces of IA3[J]. AIMS Biophysics, 2018, 5(3): 166-181. doi: 10.3934/biophy.2018.3.166







 DownLoad:
DownLoad: