Gamma radiation at therapeutic doses can cause conformation changes in proteins and consequently damage cells/tissues associated with the initiation of several pathological disorders. In this study, serum albumin, the most abundant protein in plasma, was chosen as the protein sample.
Bovine serum albumin (BSA) was exposed to gamma radiation at a therapeutic dose (3 Gy) in the absence and presence of Ceria nanoparticles (CNPs) and flower-like Fe3O4 microparticles (FIOMPs). The conformational changes in BSA including primary, secondary, tertiary structures were then studied by UV-Vis, circular dichroism (CD), and fluorescence spectroscopy, respectively.
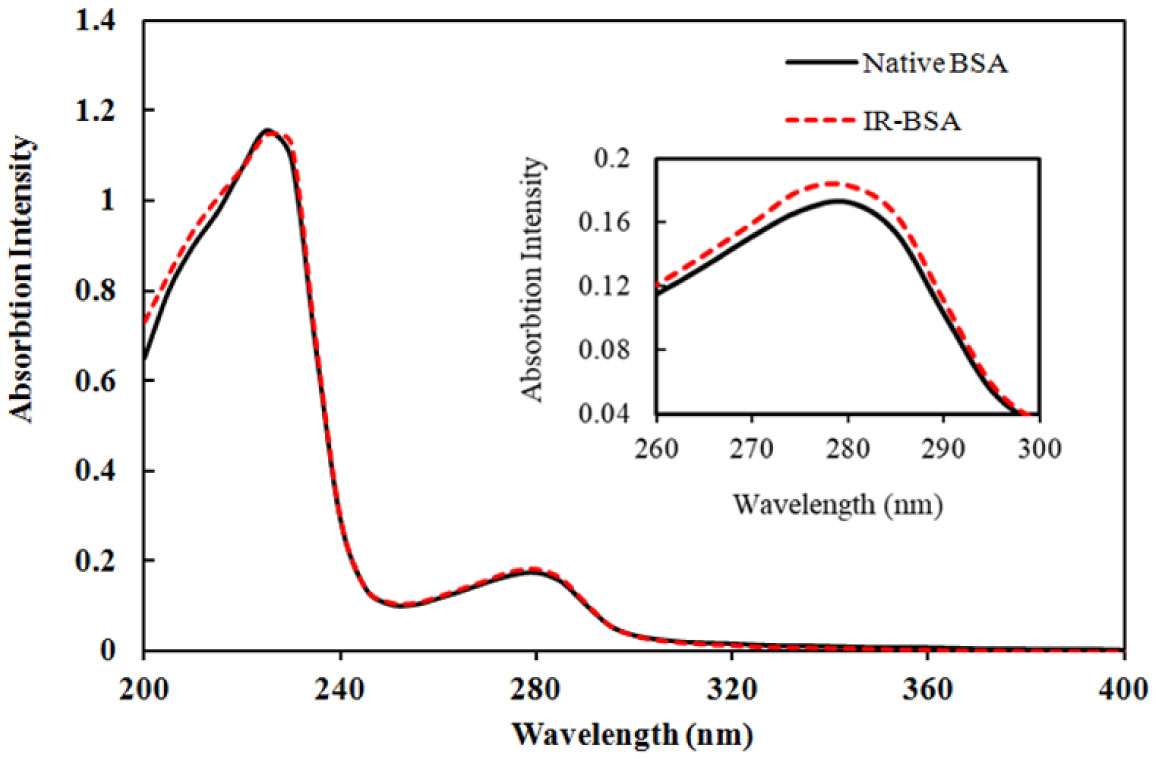
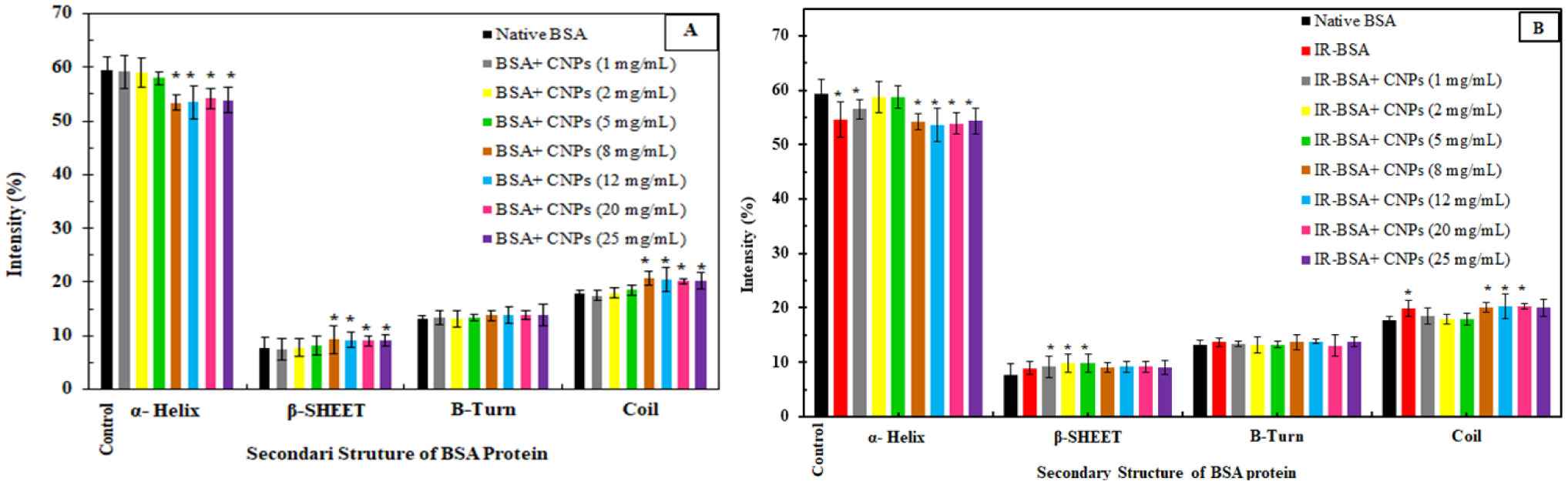
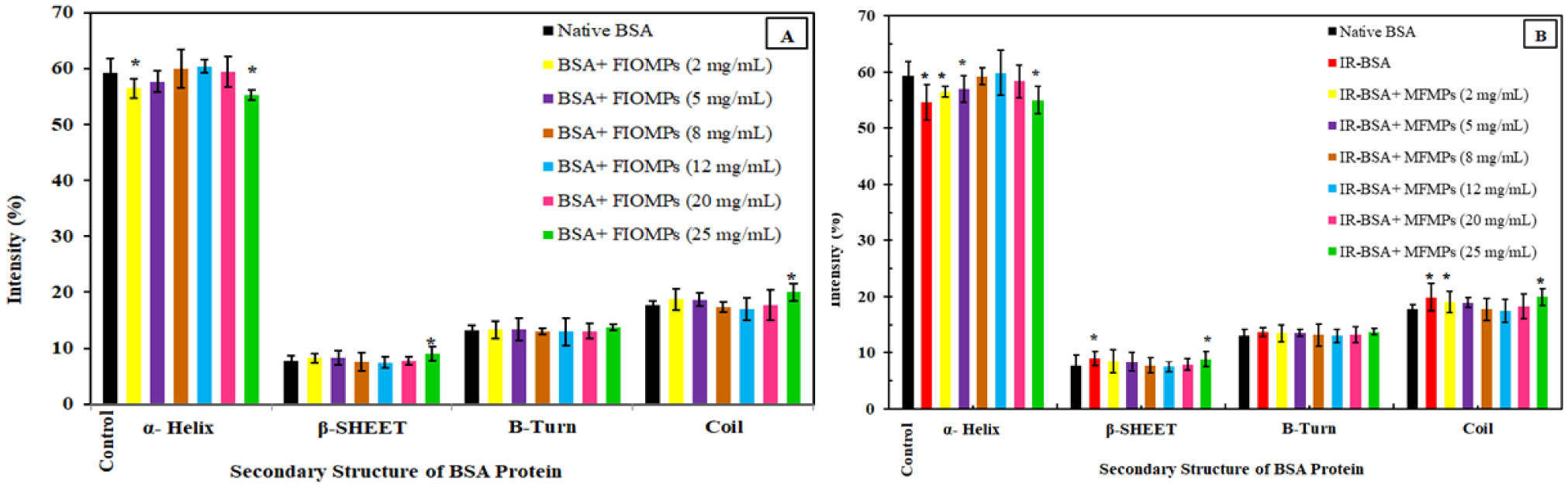
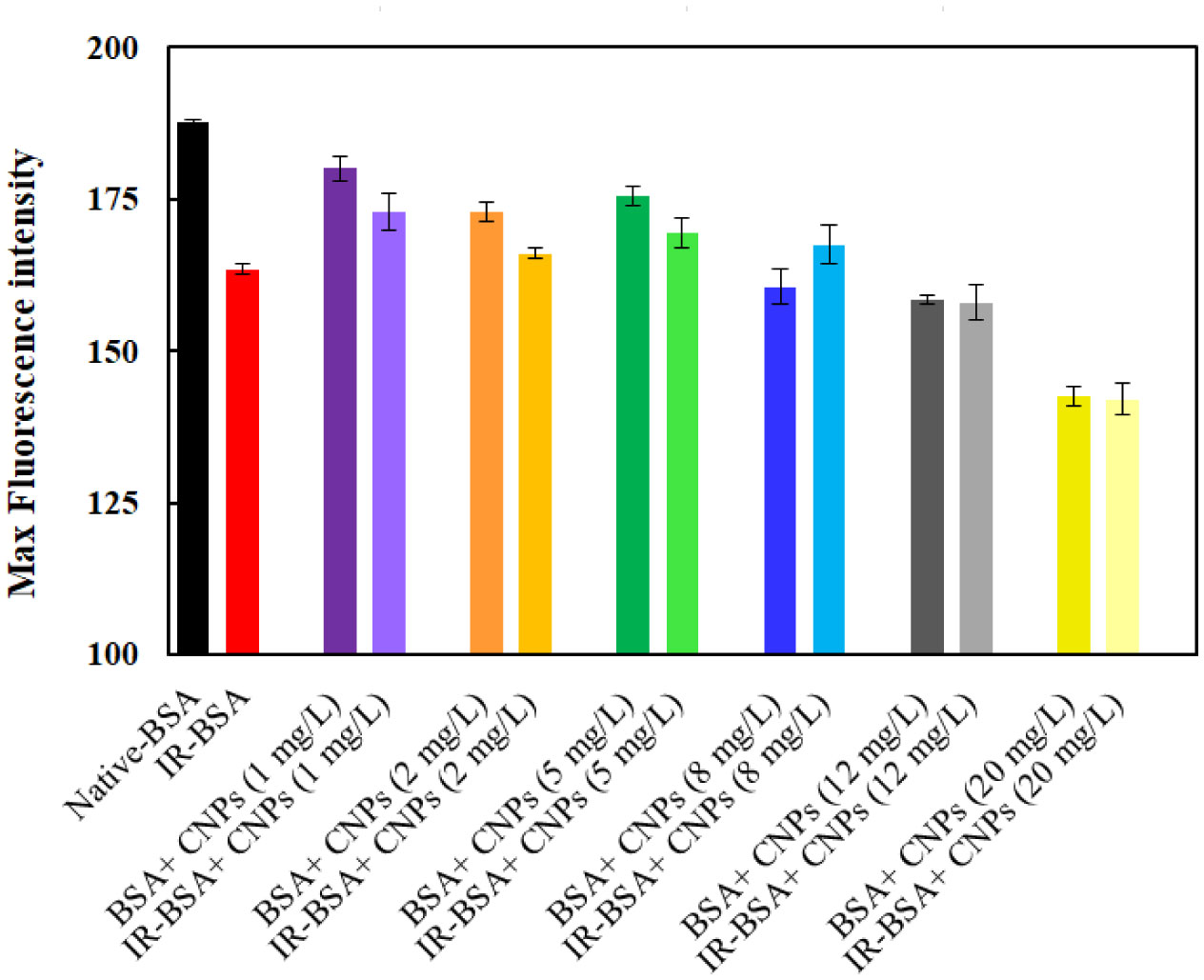
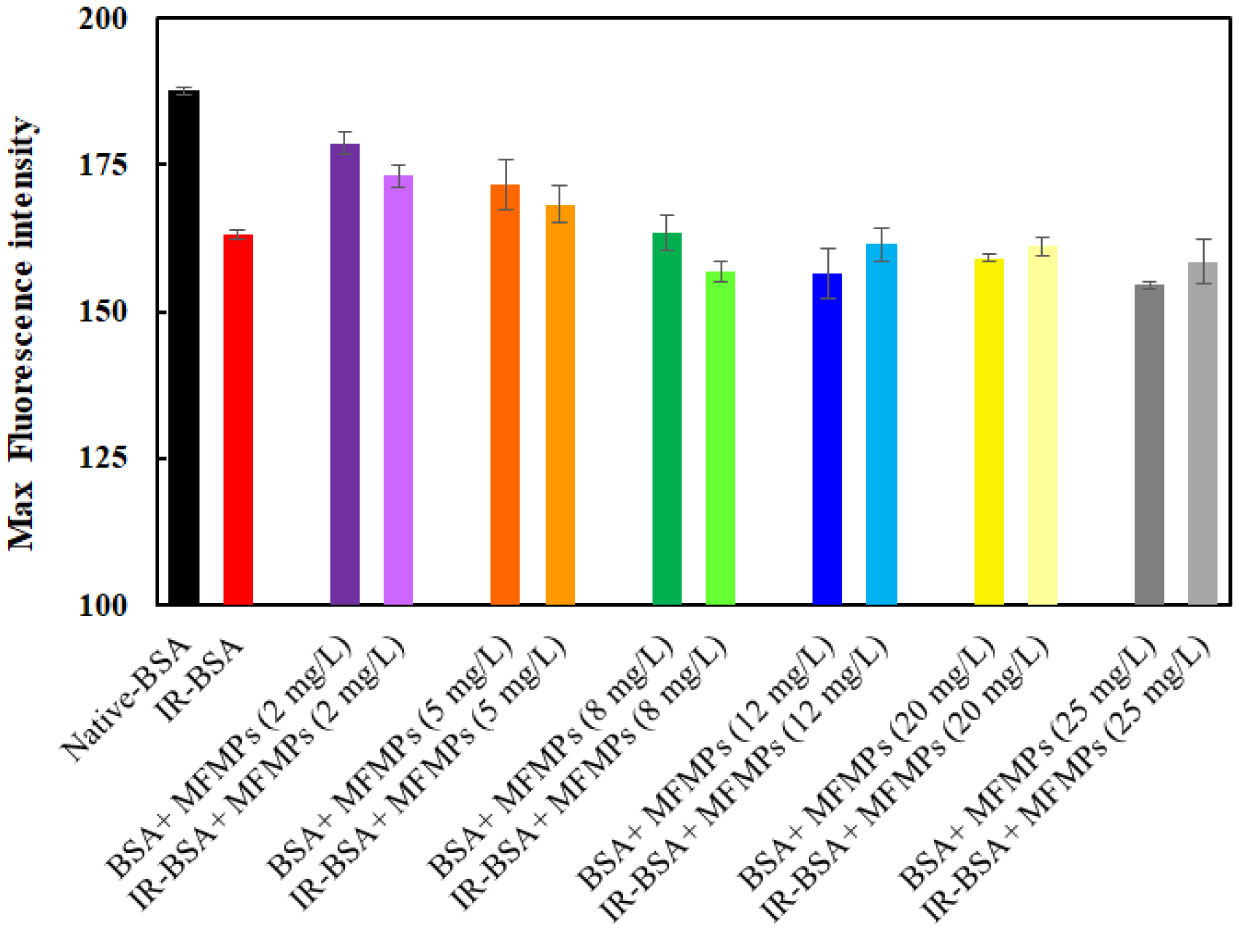
The primary structure of gamma-irradiated BSA (IR-BSA) was conserved, whereas the secondary and tertiary structures were considerably changed. IR-BSA showed α-helix to β-sheet and random coil structure transition along with reduced fluorescence emission intensity compared to non-irradiated native BSA. Both CNPs and FIOMPs could inhibit the secondary and tertiary structural changes in IR-BSA by scavenging the reactive oxygen species produced during the radiolysis of water.
The radioprotective property of CNPs arises from enzyme mimetic activities (catalase, superoxide dismutase, and peroxidase) and their antioxidant capability against hydroxyl radicals. In case of FIOMPs, the radioprotective property is attributed to catalase mimetic activity (CAT), and a porous structure leading to increased ROS recombination with each other in the same radiolytic track, and subsequently decreased encounters with BSA. The latter mechanism of restricting ROS migration seems to be more dominant for FIOMPs. Both CNPs/FIOMPs themselves at low concentrations do not show a significant effect on the native protein conformation. These findings indicate that the proposed NPs/MPs can be good candidates for developing strong nano-radioprotectors.
Citation: Mostean Bahreinipour, Hajar Zarei, Fariba Dashtestani, Jamal Rashidiani, Khadijeh Eskandari, Seyed Ali Moussavi Zarandi, Susan Kabudanian Ardestani, Hiroshi Watabe. Radioprotective effect of nanoceria and magnetic flower-like iron oxide microparticles on gamma radiation-induced damage in BSA protein[J]. AIMS Biophysics, 2021, 8(2): 124-142. doi: 10.3934/biophy.2021010
Gamma radiation at therapeutic doses can cause conformation changes in proteins and consequently damage cells/tissues associated with the initiation of several pathological disorders. In this study, serum albumin, the most abundant protein in plasma, was chosen as the protein sample.
Bovine serum albumin (BSA) was exposed to gamma radiation at a therapeutic dose (3 Gy) in the absence and presence of Ceria nanoparticles (CNPs) and flower-like Fe3O4 microparticles (FIOMPs). The conformational changes in BSA including primary, secondary, tertiary structures were then studied by UV-Vis, circular dichroism (CD), and fluorescence spectroscopy, respectively.
The primary structure of gamma-irradiated BSA (IR-BSA) was conserved, whereas the secondary and tertiary structures were considerably changed. IR-BSA showed α-helix to β-sheet and random coil structure transition along with reduced fluorescence emission intensity compared to non-irradiated native BSA. Both CNPs and FIOMPs could inhibit the secondary and tertiary structural changes in IR-BSA by scavenging the reactive oxygen species produced during the radiolysis of water.
The radioprotective property of CNPs arises from enzyme mimetic activities (catalase, superoxide dismutase, and peroxidase) and their antioxidant capability against hydroxyl radicals. In case of FIOMPs, the radioprotective property is attributed to catalase mimetic activity (CAT), and a porous structure leading to increased ROS recombination with each other in the same radiolytic track, and subsequently decreased encounters with BSA. The latter mechanism of restricting ROS migration seems to be more dominant for FIOMPs. Both CNPs/FIOMPs themselves at low concentrations do not show a significant effect on the native protein conformation. These findings indicate that the proposed NPs/MPs can be good candidates for developing strong nano-radioprotectors.

| [1] | Colon J, Hsieh N, Ferguson A, et al. (2010) Cerium oxide nanoparticles protect gastrointestinal epithelium from radiation-induced damage by reduction of reactive oxygen species and upregulation of superoxide dismutase. Biol Med 6: 698-705. |
| [2] | Kwatra D, Venugopal A, Anant S (2013) Nanoparticles in radiation therapy: a summary of various approaches to enhance radiosensitization in cancer. Transl Cancer Res 2: 330-342. |
| [3] | Madhu LN, Kumari NS (2014) Radioprotective effect of sulphydryl group containing triazole derivative to modulate the radiation-induced clastogenic effects. Res in Pharm Sci 9: 23-29. |
| [4] | Borek C (2004) Antioxidants and radiation therapy. J Nutr 134: 3207S-3209S. |
| [5] | Abdou MI, Abbas OA (2009) Evaluation of diphenyl dimethyl bicarboxylate (DDB) as a probable hepato-protector in rats against whole body gamma irradiation. Biosci Res 6: 01-11. |
| [6] | Charbonneau D, Beauregard M, Tajmir-Riahi HA (2009) Structural analysis of human serum albumin complexes with cationic lipids. J Phys Chem B 113: 1777-1784. |
| [7] | Biswas S, Das R, Banerjee ER (2017) Role of free radicals in human inflammatory diseases. AIMS Biophys 4: 596-614. |
| [8] | Shen L, Tang CH (2012) Microfluidization as a potential technique to modify surface properties of soy protein isolate. Food Res Int 48: 108-118. |
| [9] | Chen M, Liu Y, Cao H, et al. (2015) The secondary and aggregation structural changes of BSA induced by trivalent chromium: A biophysical study. J Lumin 158: 116-124. |
| [10] | Yang H, Liu Q, Zhao L, et al. (2014) Fluorescence spectroscopic studies on the interaction of oleanolic acid and its triterpenoid saponins derivatives with two serum albumins. J Solution Chem 43: 774-786. |
| [11] | Gelamo EL, Silva C, Imasato H, et al. (2002) Interaction of bovine (BSA) and human (HSA) serum albumins with ionic surfactants: spectroscopy and modelling. BBA-Protein Struct Mol Enzymol 1594: 84-99. |
| [12] | Gaber MH (2005) Effect of γ-irradiation on the molecular properties of bovine serum albumin. J Biosci Bioeng 100: 203-206. |
| [13] | Khorolskyi OV, Malomuzh NP (2020) Macromolecular sizes of serum albumins in its aqueous solutions. AIMS Biophys 7: 219-235. |
| [14] | Chudzik M, Maciążek-Jurczyk M, Pawełczak B, et al. (2016) Spectroscopic studies on the molecular ageing of serum albumin. Molecules 22: 34. |
| [15] | Schuessler H, Schilling K (1984) Oxygen effect in the radiolysis of proteins: Part 2 bovine serum albumin. Inter J Radiat Biol Relat Stud Phys, Chem Med 45: 267-281. |
| [16] | Hu X, Song W, Li W, et al. (2016) Effects of γ-irradiation on the molecular structures and functions of human serum albumin. J Biochem Mol Toxic 30: 525-532. |
| [17] | Zarei H, Bahreinipour M, Eskandari K, et al. (2017) Spectroscopic study of gamma irradiation effect on the molecular structure of bovine serum albumin. Vacuum 136: 91-96. |
| [18] | Noroozi M, Angerson WJ, Lean ME (1988) Effects of flavonoids and vitamin C on oxidative DNA damage to human lymphocytes. Am J Clin Nutr 67: 1210-1218. |
| [19] | Kumar B, Jha MN, Cole WC, et al. (2002) D-alpha-tocopheryl succinate (vitamin E) enhances radiation-induced chromosomal damage levels in human cancer cells, but reduces it in normal cells. J Am Coll Nutr 21: 339-343. |
| [20] | Mishra K, Ojha H, Kallepalli S, et al. (2014) Protective effect of ferulic acid on ionizing radiation induced damage in bovine serum albumin. Int J Radiat Res 12: 113. |
| [21] | Abdel Fattah SM, Fahim TM, El-Fatih NM (2011) Prophylactic role of combined treatment with wheat germ oil and ginseng against radiation injury in male rats. Egypt J Hosp Med 45: 403-415. |
| [22] | Verma S, Gupta ML, Dutta A, et al. (2010) Modulation of ionizing radiation induced oxidative imbalance by semi-fractionated extract of Piper betel: an in vitro and in vivo assessment. Oxid Med Cell Longev 3: 44-52. |
| [23] | Ferreira CA, Ni D, Rosenkrans ZT, et al. (2018) Scavenging of reactive oxygen and nitrogen species with nanomaterials. Nano Res 11: 4955-4984. |
| [24] | Singh S (2019) Nanomaterials exhibiting enzyme-like properties (Nanozymes): Current advances and future perspectives. Front Chem 7: 46. |
| [25] | Colon J, Herrera L, Smith J, et al. (2009) Protection from radiation-induced pneumonitis using cerium oxide nanoparticles. Nanomed: Nanotechnol 5: 225-231. |
| [26] | Tarnuzzer RW, Colon J, Patil S, et al. (2005) Vacancy engineered ceria nanostructures for protection from radiation-induced cellular damage. Nano Lett 5: 2573-2577. |
| [27] | Chen Z, Yin JJ, Zhou YT, et al. (2012) Dual enzyme-like activities of iron oxide nanoparticles and their implication for diminishing cytotoxicity. ACS Nano 6: 4001-4012. |
| [28] | Wei H, Wang E (2008) Fe3O4 magnetic nanoparticles as peroxidase mimetics and their applications in H2O2 and glucose detection. Anal Chem 80: 2250-2254. |
| [29] | Liu X, Wei W, Yuan Q, et al. (2012) Apoferritin-CeO2 nano-truffle that has excellent artificial redox enzyme activity. Chem Commun 48: 3155-3157. |
| [30] | Zhong LS, Hu JS, Liang HP, et al. (2006) Self-Assembled 3D flowerlike iron oxide nanostructures and their application in water treatment. Adv Mater 18: 2426-2431. |
| [31] | Chi Z, Liu R (2011) Phenotypic characterization of the binding of tetracycline to human serum albumin. Biomacromolecules 12: 203-209. |
| [32] | Pignataro MF, Herrera MG, Dodero VI (2020) Evaluation of peptide/protein self-assembly and aggregation by spectroscopic methods. Molecules 25: 4854. |
| [33] | Le Caër S (2011) Water radiolysis: influence of oxide surfaces on H2 production under ionizing radiation. Water 3: 235-253. |
| [34] | Uygun-Sarıbay M, Ergun E, Kalaycı Y, et al. (2017) The secondary structure of proteins in liquid, frozen, and dried egg-white samples: Effect of gamma irradiation treatment. Int J Food Prop 20: 1195-1203. |
| [35] | Zolghadri S, Saboury AA, Amin E, et al. (2010) A spectroscopic study on the interaction between ferric oxide nanoparticles and human hemoglobin. J Iran Chem Soc 7: S145-S153. |
| [36] | Hollóczki O, Gehrke S (2019) Nanoplastics can change the secondary structure of proteins. Sci Rep 9: 1-7. |
| [37] | Gopinath PM, Saranya V, Vijayakumar S, et al. (2019) Assessment on interactive prospectives of nanoplastics with plasma proteins and the toxicological impacts of virgin, coronated and environmentally released-nanoplastics. Sci Rep 9: 8860. |
| [38] | Ray D, Paul BK, Guchhait N (2012) Effect of biological confinement on the photophysics and dynamics of a proton-transfer phototautomer: an exploration of excitation and emission wavelength-dependent photophysics of the protein-bound drug. Phys Chem Chem Phys 14: 12182-12192. |
| [39] | Wang Y, Zhang H, Kang Y, et al. (2016) Effects of perfluorooctane sulfonate on the conformation and activity of bovine serum albumin. J Photoch Photobio B: Biol 159: 66-73. |
| [40] | Pal S, Pyne P, Samanta N, et al. (2020) Thermal stability modulation of the native and chemically-unfolded state of bovine serum albumin by amino acids. Phys Chem Chem Phys 22: 179-188. |
| [41] | Pant MP, Mariam J, Joshi A, et al. (2014) UV radiation sensitivity of bovine serum albumin bound to silver nanoparticles. J Radiat Res Appl Sci 7: 399-405. |
| [42] | Lu JQ, Jin F, Sun TQ, et al. (2007) Multi-spectroscopic study on interaction of bovine serum albumin with lomefloxacin–copper (II) complex. Int J Biol Macromol 40: 299-304. |
| [43] | Borowska M, Mylkie K, Nowak P, et al. (2020) Testing for ketoprofen binding to HSA coated magnetic nanoparticles under normal conditions and after oxidative stress. Molecules 25: 1945. |
| [44] | Nasir I, Lundqvist M, Cabaleiro-Lago C (2015) Size and surface chemistry of nanoparticles lead to a variant behavior in the unfolding dynamics of human carbonic anhydrase. Nanoscale 7: 17504-17515. |
| [45] | Gagner JE, Lopez MD, Dordick JS, et al. (2011) Effect of gold nanoparticle morphology on adsorbed protein structure and function. Biomaterials 32: 7241-7252. |
| [46] | Wu X, Narsimhan G (2008) Effect of surface concentration on secondary and tertiary conformational changes of lysozyme adsorbed on silica nanoparticles. BBA-Proteins Proteom 1784: 1694-1701. |
| [47] | Chetty R, Singh M (2020) In-vitro interaction of cerium oxide nanoparticles with hemoglobin, insulin, and dsDNA at 310.15 K: Physicochemical, spectroscopic and in-silico study. Int J Biol Macromol 156: 1022-1044. |
| [48] | Shemetov AA, Nabiev I, Sukhanova A (2012) Molecular interaction of proteins and peptides with nanoparticles. ACS Nano 6: 4585-4602. |
| [49] | Litvinov RI, Faizullin DA, Zuev YF, et al. (2012) The α-helix to β-sheet transition in stretched and compressed hydrated fibrin clots. Biophys J 103: 1020-1027. |
| [50] | Wei YS, Lin SY, Wang SL, et al. (2003) Fourier transform IR attenuated total reflectance spectroscopy studies of cysteine-induced changes in secondary conformations of bovine serum albumin after UV-B irradiation. Biopolymers 72: 345-351. |
| [51] | Salvia A, Carrupta PA, Tillement JP, et al. (2001) Structural damage to proteins caused by free radicals: asessment, protection by antioxidants, and influence of protein binding. Biochem Pharmacol 61: 1237-1242. |
| [52] | Gao L, Fan K, Yan X (2020) Iron oxide nanozyme: A multifunctional enzyme mimetics for biomedical application. Nanozymology Singapore: Springer, 105-140. |
| [53] | Singh S (2019) Nanomaterials exhibiting enzyme-like properties (Nanozymes): Current advances and future perspectives. Front Chem 7: 46. |
| [54] | Korsvik C, Patil S, Seal S, et al. (2007) Superoxide dismutase mimetic properties exhibited by vacancy engineered ceria nanoparticles. Chem Commun 10: 1056-1058. |
| [55] | Foley S, Rotureau P, Pin S, et al. (2005) Radiolysis of confined water: production and reactivity of hydroxyl radicals. Angew Chem Int Edit 44: 110-112. |
| [56] | Rotureau P, Renault JP, Lebeau B, et al. (2005) Radiolysis of confined water: molecular hydrogen formation. ChemPhysChem 6: 1316-1323. |
| [57] | Singh R, Singh S (2015) Role of phosphate on stability and catalase mimetic activity of cerium oxide nanoparticles. Colloid Surface B 132: 78-84. |
| [58] | Schlick S, Danilczuk M, Drews AR, et al. (2016) Scavenging of hydroxyl radicals by ceria nanoparticles: effect of particle size and concentration. J Phys Chem C 120: 6885-6890. |
| [59] | Xue Y, Luan Q, Yang D, et al. (2011) Direct evidence for hydroxyl radical scavenging activity of cerium oxide nanoparticles. J Phys Chem C 11: 4433-4438. |
| [60] | Peng FF, Zhang Y, Gu N (2008) Size-dependent peroxidase-like catalytic activity of Fe3O4 nanoparticles. Chinese Chem Lett 19: 730-733. |
| [61] | Patil US, Adireddy S, Jaiswal A, et al. (2015) In vitro/in vivo toxicity evaluation and quantification of iron oxide nanoparticles. Int J Mol Sci 16: 24417. |
| [62] | Kumari M, Singh SP, Chinde S, et al. (2014) Toxicity study of cerium oxide nanoparticles in human neuroblastoma cells. Int J Toxicol 33: 86-97. |
| [63] | Demokritou P, Gass S, Pyrgiotakis G, et al. (2013) An in vivo and in vitro toxicological characterization of realistic nanoscale CeO2 inhalation exposures. Nanotoxicology 7: 1338-1350. |
| [64] | Gagnon J, Fromm KM (2015) Toxicity and protective effects of cerium oxide nanoparticles (nanoceria) depending on their preparation method, particle size, cell type, and exposure route. Eur J Inorg Chem 27: 4510-4517. |
| [65] | Leung YH, Yung MMN, Ng AMC, et al. (2015) Toxicity of CeO2 nanoparticles–The effect of nanoparticle properties. J Photochem Photobiol B: Biol 145: 48-59. |
 biophy-08-02-010-s001.pdf biophy-08-02-010-s001.pdf |
 |

Figures(6) / Tables(2)

Mostean Bahreinipour, Hajar Zarei, Fariba Dashtestani, Jamal Rashidiani, Khadijeh Eskandari, Seyed Ali Moussavi Zarandi, Susan Kabudanian Ardestani, Hiroshi Watabe. Radioprotective effect of nanoceria and magnetic flower-like iron oxide microparticles on gamma radiation-induced damage in BSA protein[J]. AIMS Biophysics, 2021, 8(2): 124-142. doi: 10.3934/biophy.2021010







 DownLoad:
DownLoad: