Citation: Satoru Ito, Kishio Furuya, Masahiro Sokabe, Yoshinori Hasegawa. Cellular ATP release in the lung and airway[J]. AIMS Biophysics, 2016, 3(4): 571-584. doi: 10.3934/biophy.2016.4.571

| [1] |
Burnstock G, Brouns I, Adriaensen D, et al. (2012) Purinergic signaling in the airways. Pharmacol Rev 64: 834–868. doi: 10.1124/pr.111.005389

|
| [2] |
Pelleg A, Schulman ES, Barnes PJ (2016) Extracellular ATP in obstructive airway diseases. Chest 150: 908–915. doi: 10.1016/j.chest.2016.06.045
|
| [3] |
Idzko M, Hammad H, Nimwegen MV, et al. (2007) Extracellular ATP triggers and maintains asthmatic airway inflammation by activating dendritic cells. Nat Med 13: 913–919. doi: 10.1038/nm1617
|
| [4] |
Mortaz E, Folkerts G, Nijkamp FP, et al. (2010) ATP and the pathogenesis of COPD. Eur J Pharmacol 638: 1–4. doi: 10.1016/j.ejphar.2010.04.019
|
| [5] |
Riteau N, Gasse P, Fauconnier L, et al. (2010) Extracellular ATP is a danger signal activating P2X7 receptor in lung inflammation and fibrosis. Am J Respir Crit Care Med 182: 774–783. doi: 10.1164/rccm.201003-0359OC
|
| [6] |
Esther CR, Alexis NE, Clas ML, et al. (2008) Extracellular purines are biomarkers of neutrophilic airway inflammation. Eur Respir J 31: 949–956. doi: 10.1183/09031936.00089807
|
| [7] |
Lazarowski ER, Boucher RC, Harden TK (2000) Constitutive release of ATP and evidence for major contribution of ecto-nucleotide pyrophosphatase and nucleoside diphosphokinase to extracellular nucleotide concentrations. J Biol Chem 275: 31061–31068. doi: 10.1074/jbc.M003255200
|
| [8] |
Praetorius HA, Leipziger J (2009) ATP release from non-excitable cells. Purinergic Signal 5: 433–446. doi: 10.1007/s11302-009-9146-2
|
| [9] |
Adriaensen D, Timmermans JP (2004) Purinergic signalling in the lung: important in asthma and COPD? Curr Opin Pharmacol 4: 207–214. doi: 10.1016/j.coph.2004.01.010
|
| [10] |
Burnstock G (2006) Historical review: ATP as a neurotransmitter. Trends Pharmacol Sci 27: 166–176. doi: 10.1016/j.tips.2006.01.005
|
| [11] | Grygorczyk R, Hanrahan JW (1997) CFTR-independent ATP release from epithelial cells triggered by mechanical stimuli. Am J Physiol 272: 1058–1066. |
| [12] |
Murata N, Ito S, Furuya K, et al. (2014) Ca2+ influx and ATP release mediated by mechanical stretch in human lung fibroblasts. Biochem Biophys Res Commun 453: 101–105. doi: 10.1016/j.bbrc.2014.09.063
|
| [13] |
Takahara N, Ito S, Furuya K, et al. (2014) Real-time imaging of ATP release induced by mechanical stretch in human airway smooth muscle cells. Am J Respir Cell Mol Biol 51: 772–782. doi: 10.1165/rcmb.2014-0008OC
|
| [14] |
Yamamoto K, Furuya K, Nakamura M, et al. (2011) Visualization of flow-induced ATP release and triggering of Ca2+ waves at caveolae in vascular endothelial cells. J Cell Sci 124: 3477–3483. doi: 10.1242/jcs.087221
|
| [15] |
Yamamoto K, Sokabe T, Matsumoto T, et al. (2006) Impaired flow-dependent control of vascular tone and remodeling in P2X4-deficient mice. Nat Med 12: 133–137. doi: 10.1038/nm1338
|
| [16] | Ralevic V, Burnstock G (1998) Receptors for purines and pyrimidines. Pharmacol Rev 50: 413–492. |
| [17] |
Knowles MR, Clarke LL, Boucher RC (1991) Activation by extracellular nucleotides of chloride secretion in the airway epithelia of patients with cystic fibrosis. N Engl J Med 325: 533–538. doi: 10.1056/NEJM199108223250802
|
| [18] |
Son M, Ito Y, Sato S, et al. (2004) Apical and basolateral ATP-induced anion secretion in polarized human airway epithelia. Am J Respir Cell Mol Biol 30: 411–419. doi: 10.1165/rcmb.2003-0183OC
|
| [19] |
Rice WR (1990) Effects of extracellular ATP on surfactant secretion. Ann N Y Acad Sci 603: 64–74. doi: 10.1111/j.1749-6632.1990.tb37662.x
|
| [20] | Button B, Okada SF, Frederick CB, et al. (2013) Mechanosensitive ATP release maintains proper mucus hydration of airways. Sci Signal 6: ra46. |
| [21] |
Button B, Picher M, Boucher RC (2007) Differential effects of cyclic and constant stress on ATP release and mucociliary transport by human airway epithelia. J Physiol 580: 577–592. doi: 10.1113/jphysiol.2006.126086
|
| [22] |
Kiefmann R, Islam MN, Lindert J, et al. (2009) Paracrine purinergic signaling determines lung endothelial nitric oxide production. Am J Physiol Lung Cell Mol Physiol 296: 901–910. doi: 10.1152/ajplung.90549.2008
|
| [23] |
Boucher RC (2007) Airway surface dehydration in cystic fibrosis: pathogenesis and therapy. Annu Rev Med 58: 157–170. doi: 10.1146/annurev.med.58.071905.105316
|
| [24] |
Karp CL, Flick LM, Park KW, et al. (2004) Defective lipoxin-mediated anti-inflammatory activity in the cystic fibrosis airway. Nat Immunol 5: 388–392. doi: 10.1038/ni1056
|
| [25] | Higgins G, Buchanan P, Perriere M, et al. (2014) Activation of P2RY11 and ATP release by lipoxin A4 restores the airway surface liquid layer and epithelial repair in cystic fibrosis. Am J Respir Cell Mol Biol 51: 178–190. |
| [26] | Ellson CD, Dunmore R, Hogaboam CM, et al. (2014) Danger-associated molecular patterns and danger signals in idiopathic pulmonary fibrosis. Am J Respir Cell Mol Biol 51: 163–168. |
| [27] |
Takai E, Tsukimoto M, Harada H, et al. (2014) Autocrine signaling via release of ATP and activation of P2X7 receptor influences motile activity of human lung cancer cells. Purinergic Signal 10: 487–497. doi: 10.1007/s11302-014-9411-x
|
| [28] |
Mortaz E, Braber S, Nazary M, et al. (2009) ATP in the pathogenesis of lung emphysema. Eur J Pharmacol 619: 92–96. doi: 10.1016/j.ejphar.2009.07.022
|
| [29] | Gerasimovskaya EV, Ahmad S, White CW, et al. (2002) Extracellular ATP is an autocrine/paracrine regulator of hypoxia-induced adventitial fibroblast growth. Signaling through extracellular signal-regulated kinase-1/2 and the Egr-1 transcription factor. J Biol Chem 277: 44638–44650. |
| [30] |
Burnstock G (2006) Pathophysiology and therapeutic potential of purinergic signaling. Pharmacol Rev 58: 58–86. doi: 10.1124/pr.58.1.5
|
| [31] |
Willart MA, Lambrecht BN (2009) The danger within: endogenous danger signals, atopy and asthma. Clin Exp Allergy 39: 12–19. doi: 10.1111/j.1365-2222.2008.03118.x
|
| [32] |
Pelleg A, Schulman ES (2002) Adenosine 5’-triphosphate axis in obstructive airway diseases. Am J Ther 9: 454–464. doi: 10.1097/00045391-200209000-00014
|
| [33] |
Perez-Zoghbi JF, Karner C, Ito S, et al. (2009) Ion channel regulation of intracellular calcium and airway smooth muscle function. Pulm Pharmacol Ther 22: 388–397. doi: 10.1016/j.pupt.2008.09.006
|
| [34] |
Bergner A, Sanderson MJ (2002) ATP stimulates Ca2+ oscillations and contraction in airway smooth muscle cells of mouse lung slices. Am J Physiol Lung Cell Mol Physiol 283: 1271–1279. doi: 10.1152/ajplung.00139.2002
|
| [35] |
Mounkaila B, Marthan R, Roux E (2005) Biphasic effect of extracellular ATP on human and rat airways is due to multiple P2 purinoceptor activation. Respir Res 6: 143. doi: 10.1186/1465-9921-6-143
|
| [36] |
Michoud MC, Tao FC, Pradhan AA, et al. (1999) Mechanisms of the potentiation by adenosine of adenosine triphosphate-induced calcium release in tracheal smooth-muscle cells. Am J Respir Cell Mol Biol 21: 30–36. doi: 10.1165/ajrcmb.21.1.3425
|
| [37] |
Michoud MC, Tolloczko B, Martin JG (1997) Effects of purine nucleotides and nucleoside on cytosolic calcium levels in rat tracheal smooth muscle cells. Am J Respir Cell Mol Biol 16: 199–205. doi: 10.1165/ajrcmb.16.2.9032128
|
| [38] |
Oguma T, Ito S, Kondo M, et al. (2007) Roles of P2X receptors and Ca2+ sensitization in extracellular adenosine triphosphate-induced hyperresponsiveness in airway smooth muscle. Clin Exp Allergy 37: 893–900. doi: 10.1111/j.1365-2222.2007.02719.x
|
| [39] |
Myrtek D, Idzko M (2007) Chemotactic activity of extracellular nucleotideson human immune cells. Purinergic Signal 3: 5–11. doi: 10.1007/s11302-006-9032-0
|
| [40] |
Ferrari D, Gambari R, Idzko M, et al. (2016) Purinergic signaling in scarring. FASEB J 30: 3–12. doi: 10.1096/fj.15-274563
|
| [41] |
Janssen LJ, Farkas L, Rahman T, et al. (2009) ATP stimulates Ca2+-waves and gene expression in cultured human pulmonary fibroblasts. Int J Biochem Cell Biol 41: 2477–2484. doi: 10.1016/j.biocel.2009.08.004
|
| [42] |
Hinz B, Phan SH, Thannickal VJ, et al. (2012) Recent developments in myofibroblast biology: paradigms for connective tissue remodeling. Am J Pathol 180: 1340–1355. doi: 10.1016/j.ajpath.2012.02.004
|
| [43] |
Tschumperlin DJ (2013) Fibroblasts and the ground they walk on. Physiology 28: 380–390. doi: 10.1152/physiol.00024.2013
|
| [44] |
Carloni A, Poletti V, Fermo L, et al. (2013) Heterogeneous distribution of mechanical stress in human lung: a mathematical approach to evaluate abnormal remodeling in IPF. J Theor Biol 332: 136–140. doi: 10.1016/j.jtbi.2013.04.038
|
| [45] |
Hinz B (2012) Mechanical aspects of lung fibrosis: a spotlight on the myofibroblast. Proc Am Thorac Soc 9: 137–147. doi: 10.1513/pats.201202-017AW
|
| [46] |
Boudreault F, Tschumperlin DJ (2010) Stretch-induced mitogen-activated protein kinase activation in lung fibroblasts is independent of receptor tyrosine kinases. Am J Respir Cell Mol Biol 43: 64–73. doi: 10.1165/rcmb.2009-0092OC
|
| [47] |
Ito S, Kume H, Naruse K, et al. (2008) A novel Ca2+ influx pathway activated by mechanical stretch in human airway smooth muscle cells. Am J Respir Cell Mol Biol 38: 407–413. doi: 10.1165/rcmb.2007-0259OC
|
| [48] |
Ito S, Suki B, Kume H, et al. (2010) Actin cytoskeleton regulates stretch-activated Ca2+ influx in human pulmonary microvascular endothelial cells. Am J Respir Cell Mol Biol 43: 26–34. doi: 10.1165/rcmb.2009-0073OC
|
| [49] |
Takada H, Furuya K, Sokabe M (2014) Mechanosensitive ATP release from hemichannels and Ca2+ influx through TRPC6 accelerate wound closure in keratinocytes. J Cell Sci 127: 4159–4171. doi: 10.1242/jcs.147314
|
| [50] |
Tschumperlin DJ, Boudreault F, Liu F (2010) Recent advances and new opportunities in lung mechanobiology. J Biomech 43: 99–107. doi: 10.1016/j.jbiomech.2009.09.015
|
| [51] |
Iwaki M, Ito S, Morioka M, et al. (2009) Mechanical stretch enhances IL-8 production in pulmonary microvascular endothelial cells. Biochem Biophys Res Commun 389: 531–536. doi: 10.1016/j.bbrc.2009.09.020
|
| [52] |
Morioka M, Parameswaran H, Naruse K, et al. (2011) Microtubule dynamics regulate cyclic stretch-induced cell alignment in human airway smooth muscle cells. PloS One 6: e26384. doi: 10.1371/journal.pone.0026384
|
| [53] |
Matthay MA, Zemans RL (2011) The acute respiratory distress syndrome: pathogenesis and treatment. Annu Rev Pathol 6: 147–163. doi: 10.1146/annurev-pathol-011110-130158
|
| [54] |
Slutsky AS, Ranieri VM (2013) Ventilator-induced lung injury. N Engl J Med 369: 2126–2136. doi: 10.1056/NEJMra1208707
|
| [55] | Ito S, Hasegawa Y (2012) Mechanical stretch and cytokine synthesis in pulmonary endothelial cells. In: Kamkin A, Kiseleva I, Mechanical Stretch and Cytokines, Springer, 165–187. |
| [56] |
Rich PB, Douillet CD, Mahler SA, et al. (2003) Adenosine triphosphate is released during injurious mechanical ventilation and contributes to lung edema. J Trauma 55: 290–297. doi: 10.1097/01.TA.0000078882.11919.AF
|
| [57] |
Junger WG (2011) Immune cell regulation by autocrine purinergic signalling. Nat Rev Immunol 11: 201–212. doi: 10.1038/nri2938
|
| [58] | Corriden R, Insel PA (2010) Basal release of ATP: an autocrine-paracrine mechanism for cell regulation. Sci Signal 3: re1. |
| [59] |
Lazarowski ER (2012) Vesicular and conductive mechanisms of nucleotide release. Purinergic Signal 8: 359–373. doi: 10.1007/s11302-012-9304-9
|
| [60] |
Bodin P, Bailey D, Burnstock G (1991) Increased flow-induced ATP release from isolated vascular endothelial cells but not smooth muscle cells. Br J Pharmacol 103: 1203–1205. doi: 10.1111/j.1476-5381.1991.tb12324.x
|
| [61] |
Yamamoto K, Sokabe T, Ohura N, et al. (2003) Endogenously released ATP mediates shear stress-induced Ca2+ influx into pulmonary artery endothelial cells. Am J Physiol Heart Circ Physiol 285: H793–803. doi: 10.1152/ajpheart.01155.2002
|
| [62] |
Ransford GA, Fregien N, Qiu F, et al. (2009) Pannexin 1 contributes to ATP release in airway epithelia. Am J Respir Cell Mol Biol 41: 525–534. doi: 10.1165/rcmb.2008-0367OC
|
| [63] |
Ito Y, Son M, Sato S, et al. (2004) ATP release triggered by activation of the Ca2+-activated K+ channel in human airway Calu-3 cells. Am J Respir Cell Mol Biol 30: 388–395. doi: 10.1165/rcmb.2003-0184OC
|
| [64] |
Okada SF, Ribeiro CM, Sesma JI, et al. (2013) Inflammation promotes airway epithelial ATP release via calcium-dependent vesicular pathways. Am J Respir Cell Mol Biol 49: 814–820. doi: 10.1165/rcmb.2012-0493OC
|
| [65] |
Grygorczyk R, Furuya K, Sokabe M (2013) Imaging and characterization of stretch-induced ATP release from alveolar A549 cells. J Physiol 591: 1195–1215. doi: 10.1113/jphysiol.2012.244145
|
| [66] | Beigi R, Kobatake E, Aizawa M, et al. (1999) Detection of local ATP release from activated platelets using cell surface-attached firefly luciferase. Am J Physiol 276: 267–278. |
| [67] |
Nakamura M, Mie M, Funabashi H, et al. (2006) Cell-surface-localized ATP detection with immobilized firefly luciferase. Anal Biochem 352: 61–67. doi: 10.1016/j.ab.2006.02.019
|
| [68] | Sorensen CE, Novak I (2001) Visualization of ATP release in pancreatic acini in response to cholinergic stimulus. Use of fluorescent probes and confocal microscopy. J Biol Chem 276: 32925–32932. |
| [69] |
Furuya K, Sokabe M, Grygorczyk R (2014) Real-time luminescence imaging of cellular ATP release. Methods 66: 330–344. doi: 10.1016/j.ymeth.2013.08.007
|
| [70] |
Corriden R, Insel PA, Junger WG (2007) A novel method using fluorescence microscopy for real-time assessment of ATP release from individual cells. Am J Physiol Cell Physiol 293: 1420–1425. doi: 10.1152/ajpcell.00271.2007
|
| [71] |
Kurishita Y, Kohira T, Ojida A, et al. (2012) Organelle-localizable fluorescent chemosensors for site-specific multicolor imaging of nucleoside polyphosphate dynamics in living cells. J Am Chem Soc 134: 18779–18789. doi: 10.1021/ja308754g
|
| [72] |
Ojida A, Takashima I, Kohira T, et al. (2008) Turn-on fluorescence sensing of nucleoside polyphosphates using a xanthene-based Zn (II) complex chemosensor. J Am Chem Soc 130: 12095–12101. doi: 10.1021/ja803262w
|
| [73] |
Ledderose C, Bao Y, Zhang J, et al. (2015) Novel method for real-time monitoring of ATP release reveals multiple phases of autocrine purinergic signalling during immune cell activation. Acta Physiol (Oxf) 213: 334–345. doi: 10.1111/apha.12435
|
| [74] |
Furuya K, Tan JJ, Boudreault F, et al. (2016) Real-time imaging of inflation-induced ATP release in the ex-vivo rat lung. Am J Physiol Lung Cell Mol Physiol 311: L956–L969. doi: 10.1152/ajplung.00425.2015
|
| [75] | Fitz JG (2007) Regulation of cellular ATP release. Trans Am Clin Climatol Assoc 118: 199–208. |
| [76] |
Sabirov RZ, Okada Y (2005) ATP release via anion channels. Purinergic Signal 1: 311–328. doi: 10.1007/s11302-005-1557-0
|
| [77] |
Burnstock G (2008) Unresolved issues and controversies in purinergic signalling. J Physiol 586: 3307–3312. doi: 10.1113/jphysiol.2008.155903
|
| [78] |
Zhang Z, Chen G, Zhou W, et al. (2007) Regulated ATP release from astrocytes through lysosome exocytosis. Nat Cell Biol 9: 945–953. doi: 10.1038/ncb1620
|
| [79] |
Takai E, Tsukimoto M, Harada H, et al. (2012) Autocrine regulation of TGF-beta1-induced cell migration by exocytosis of ATP and activation of P2 receptors in human lung cancer cells. J Cell Sci 125: 5051–5060. doi: 10.1242/jcs.104976
|
| [80] |
Sawada K, Echigo N, Juge N, et al. (2008) Identification of a vesicular nucleotide transporter. Proc Natl Acad Sci USA 105: 5683–5686. doi: 10.1073/pnas.0800141105
|
| [81] |
Sudhof TC (2013) A molecular machine for neurotransmitter release: synaptotagmin and beyond. Nat Med 19: 1227–1231. doi: 10.1038/nm.3338
|
| [82] |
Jahn R, Fasshauer D (2012) Molecular machines governing exocytosis of synaptic vesicles. Nature 490: 201–207. doi: 10.1038/nature11320
|
| [83] |
Sesma JI, Kreda SM, Okada SF, et al. (2013) Vesicular nucleotide transporter regulates the nucleotide content in airway epithelial mucin granules. Am J Physiol Cell Physiol 304: C976–984. doi: 10.1152/ajpcell.00371.2012
|
| [84] |
Raposo G, Stoorvogel W (2013) Extracellular vesicles: exosomes, microvesicles and friends. J Cell Biol 200: 373–383. doi: 10.1083/jcb.201211138
|
| [85] |
Fujita Y, Kosaka N, Araya J, et al. (2015) Extracellular vesicles in lung microenvironment and pathogenesis. Trends Mol Med 21: 533–542. doi: 10.1016/j.molmed.2015.07.004
|
| [86] |
Dubyak GR (2012) P2X7 receptor regulation of non-classical secretion from immune effector cells. Cell Microbiol 14: 1697–1706. doi: 10.1111/cmi.12001
|
| [87] |
Turturici G, Tinnirello R, Sconzo G, et al. (2014) Extracellular membrane vesicles as a mechanism of cell-to-cell communication: advantages and disadvantages. Am J Physiol Cell Physiol 306: 621–633. doi: 10.1152/ajpcell.00228.2013
|
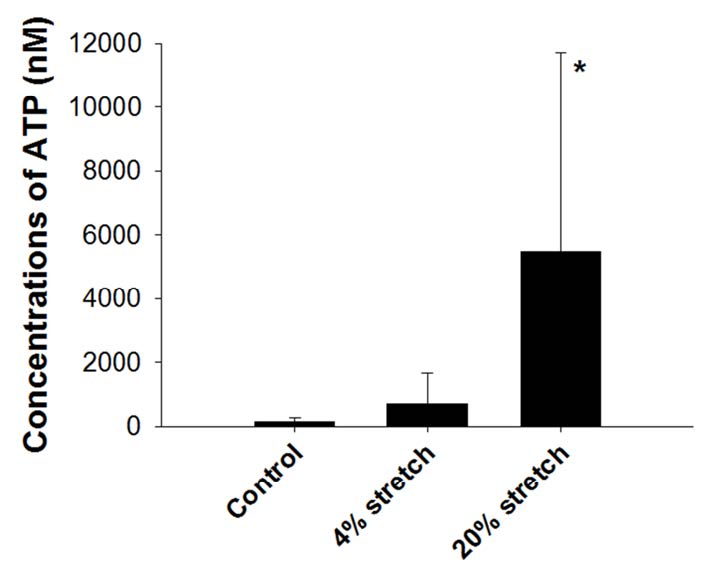
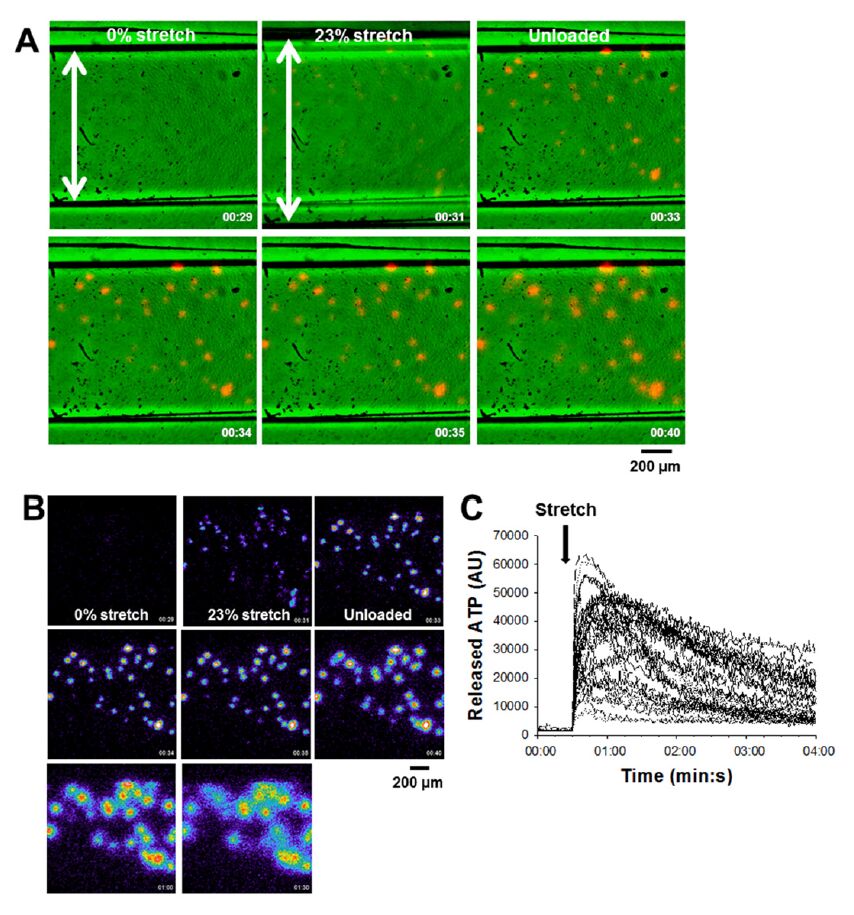
Figures(2)

Satoru Ito, Kishio Furuya, Masahiro Sokabe, Yoshinori Hasegawa. Cellular ATP release in the lung and airway[J]. AIMS Biophysics, 2016, 3(4): 571-584. doi: 10.3934/biophy.2016.4.571







 DownLoad:
DownLoad: