Citation: David E. Shoup. Diffusion-controlled reaction rates for clusters of binding sites on a cell[J]. AIMS Biophysics, 2016, 3(4): 522-528. doi: 10.3934/biophy.2016.4.522

| [1] | Berg H, Purcell E (1977) Physics of chemoreception. Biophys J 20: 193–219. |
| [2] |
Erickson J, Goldstein B, Holowka D, et al. (1987) The effect of receptor density on the forward rate constant for binding of ligands to cell surface receptors. Biophys J 52: 657–662. doi: 10.1016/S0006-3495(87)83258-7

|
| [3] | Philips R, (2013) Real receptors are not always uniformly distributed, In: Philips R, Author, Physical Biology of the Cell, London and New York: Garland Science, 536–537. |
| [4] |
Goldstein B, Wiegel F (1983) The effect of receptor clustering on diffusion-limited forward rate constants. Biophys J 43: 121–125. doi: 10.1016/S0006-3495(83)84330-6
|
| [5] | Potanin V, Verkhusha V, Belokneva O, et al. (1994) Kinetics of ligand binding to a cluster of membrane-associated receptors. Eur Biophys J 23:197–205. |
| [6] | Care B, Soula H (2011) Impact of receptor clustering on ligand binding. BMC Syst Biol 5: 48–66. |
| [7] | Shoup D, Lipari G, Szabo A (1981) Diffusion-controlled bimolecular reaction rates: The effect of rotational diffusion and orientation constraints. Biophys J 36: 697–714. |
| [8] | Traytak S (1995) Diffusion-controlled reaction rate to an active site. Chemical Physics 192: 1–7. |
| [9] | Solc K, Stockmayer W (1973) Kinetics of diffusion-controlled reaction between chemically asymmetric molecules. II. Approximate steady state solution. Int J Chem Kinet 5: 733–752. |
| [10] |
Samson R, Deutch J (1978) Diffusion-controlled reaction to a buried active site. J Chem Phys 68: 285–290. doi: 10.1063/1.435494
|
| [11] | Shoup D (2014) Diffusion-controlled reaction rates for two active sites on a sphere. BMC Biophys 7: 3–5. |
| [12] | Traytak S, Barzykin A (2007) Diffusion-controlled reaction rate on a sink with two active sites. J Chem Phys 127: 215103-1-215103-4. |
| [13] | Shoup D, Szabo A (1982) Role of diffusion in ligand binding to macromolecules and cell-bound receptors. Biophys J 40: 33–39. |
| [14] | Collins F, Kimball G (1949) Diffusion-controlled reaction rates. J Colloid Sci 4: 425–437. |
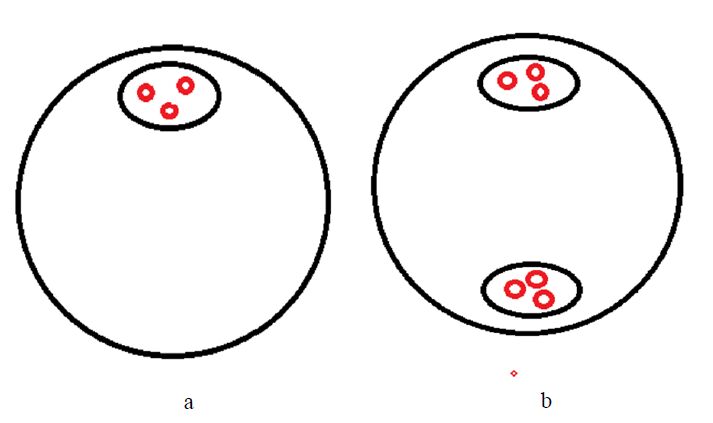
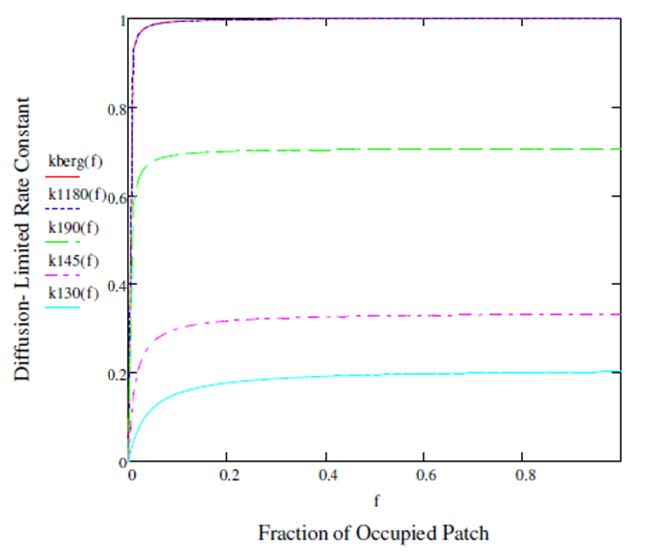
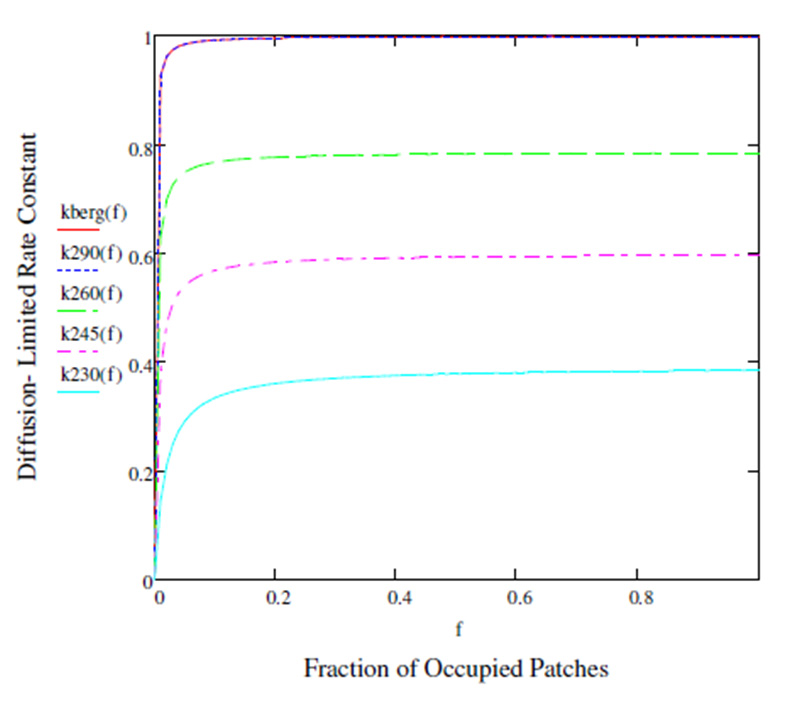
Figures(3)

David E. Shoup. Diffusion-controlled reaction rates for clusters of binding sites on a cell[J]. AIMS Biophysics, 2016, 3(4): 522-528. doi: 10.3934/biophy.2016.4.522







 DownLoad:
DownLoad: