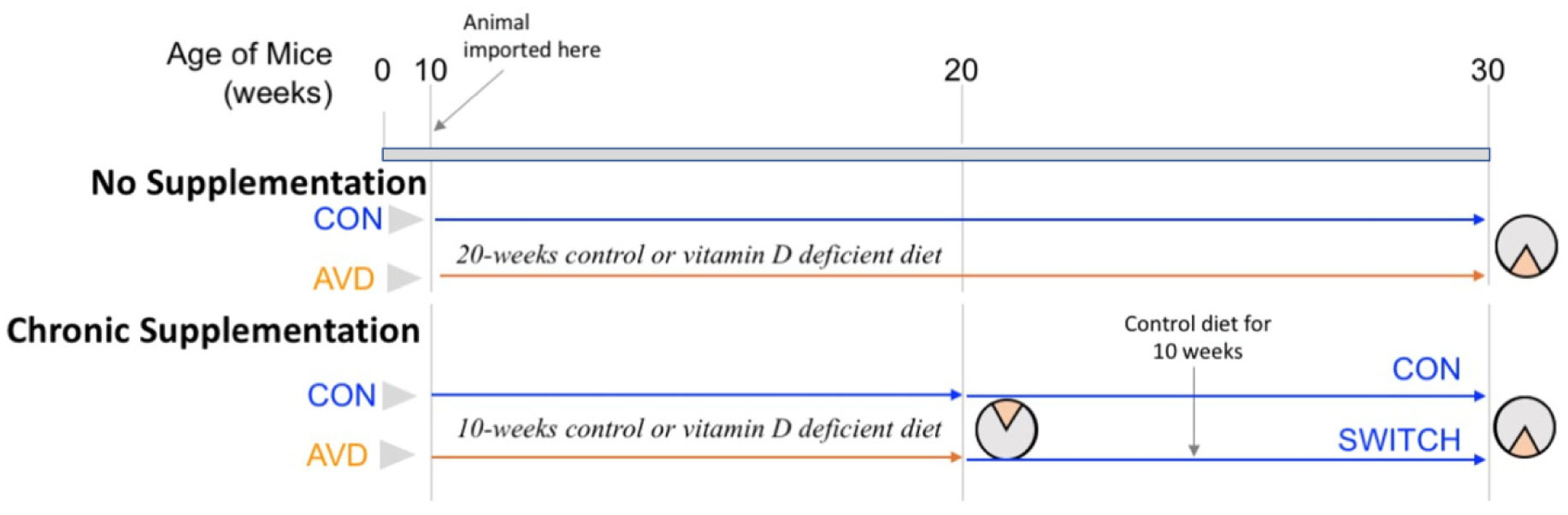
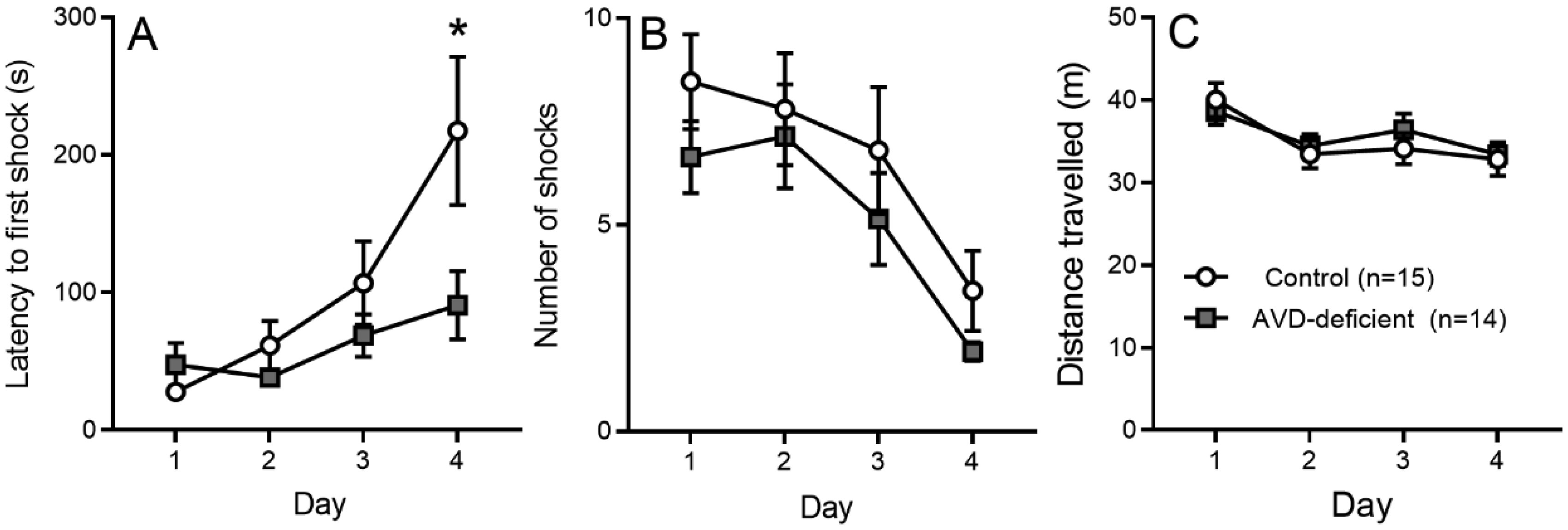
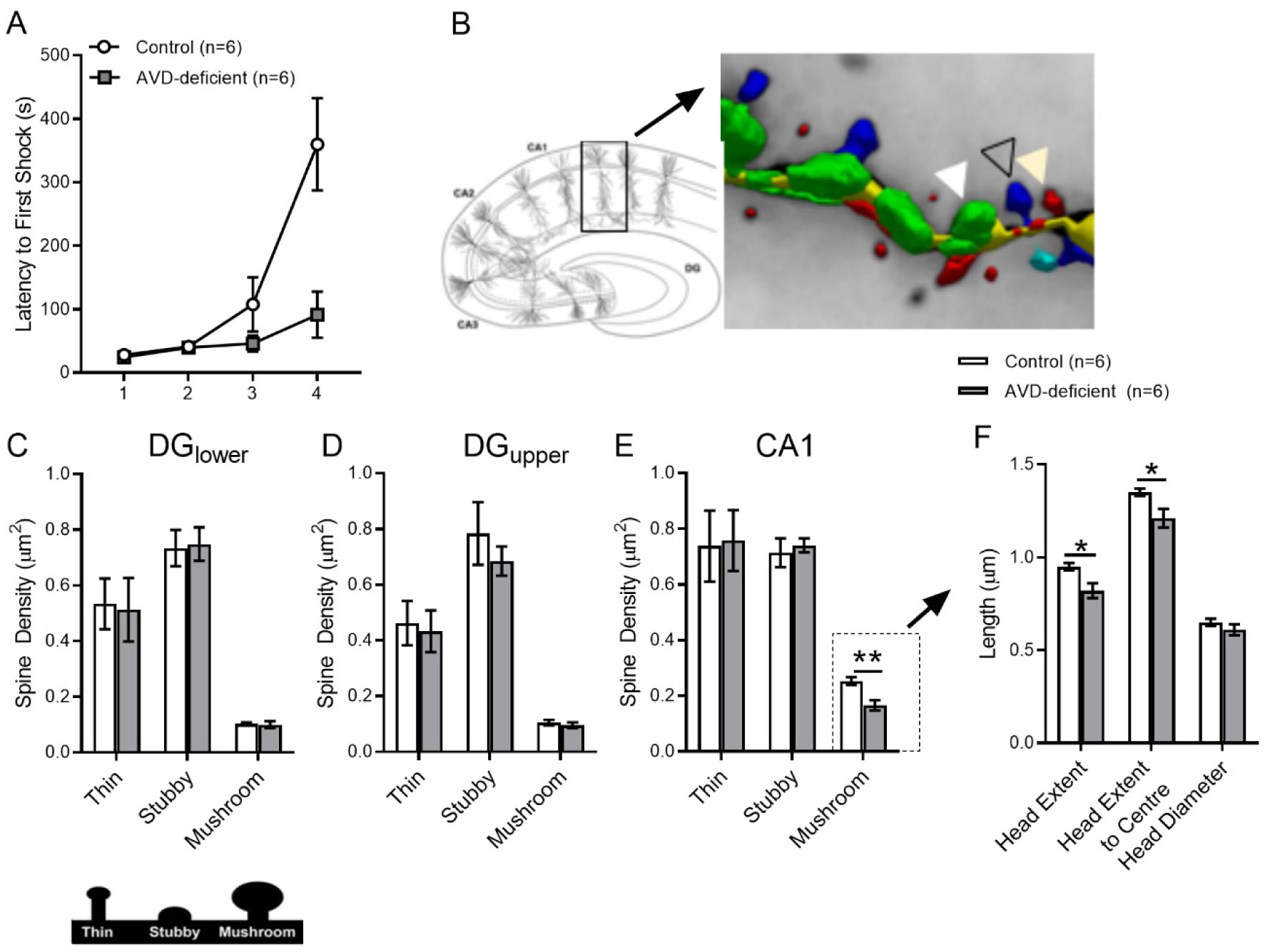
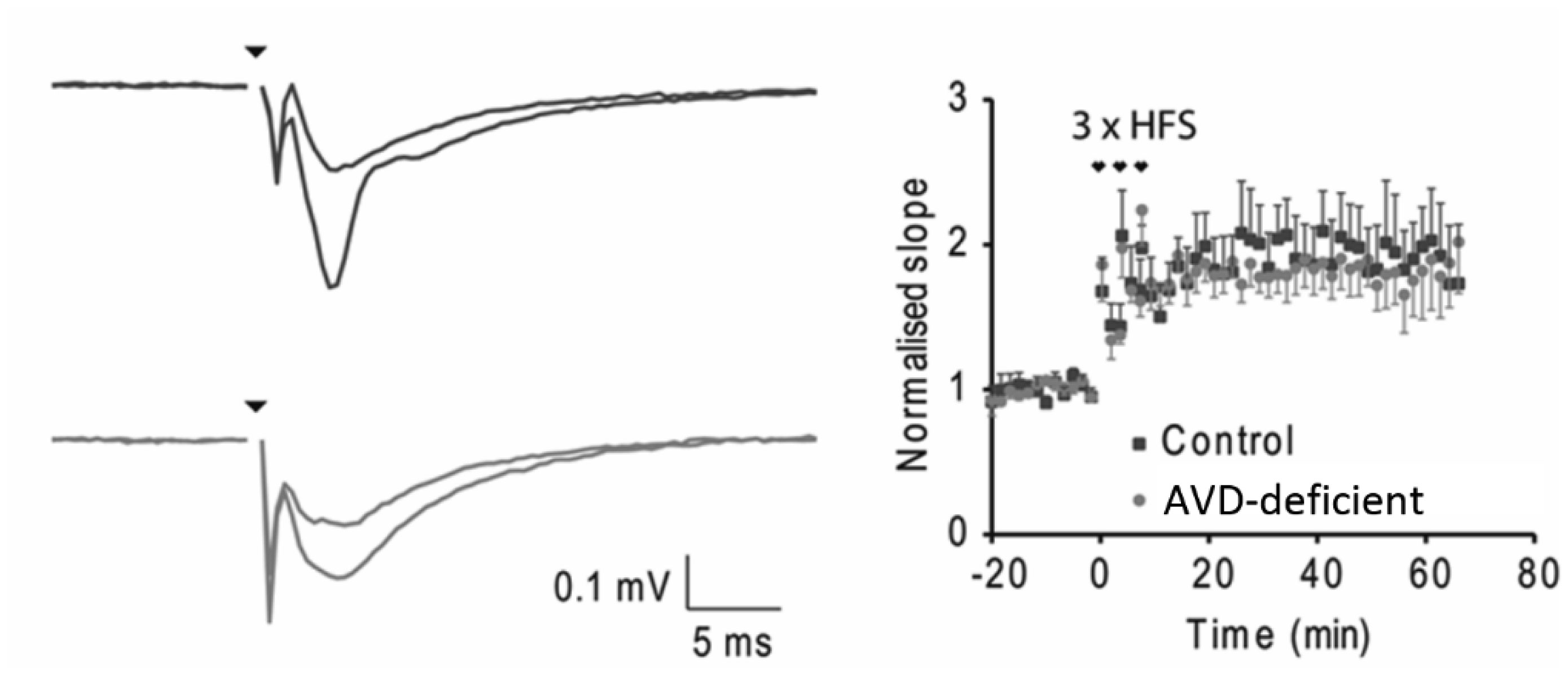
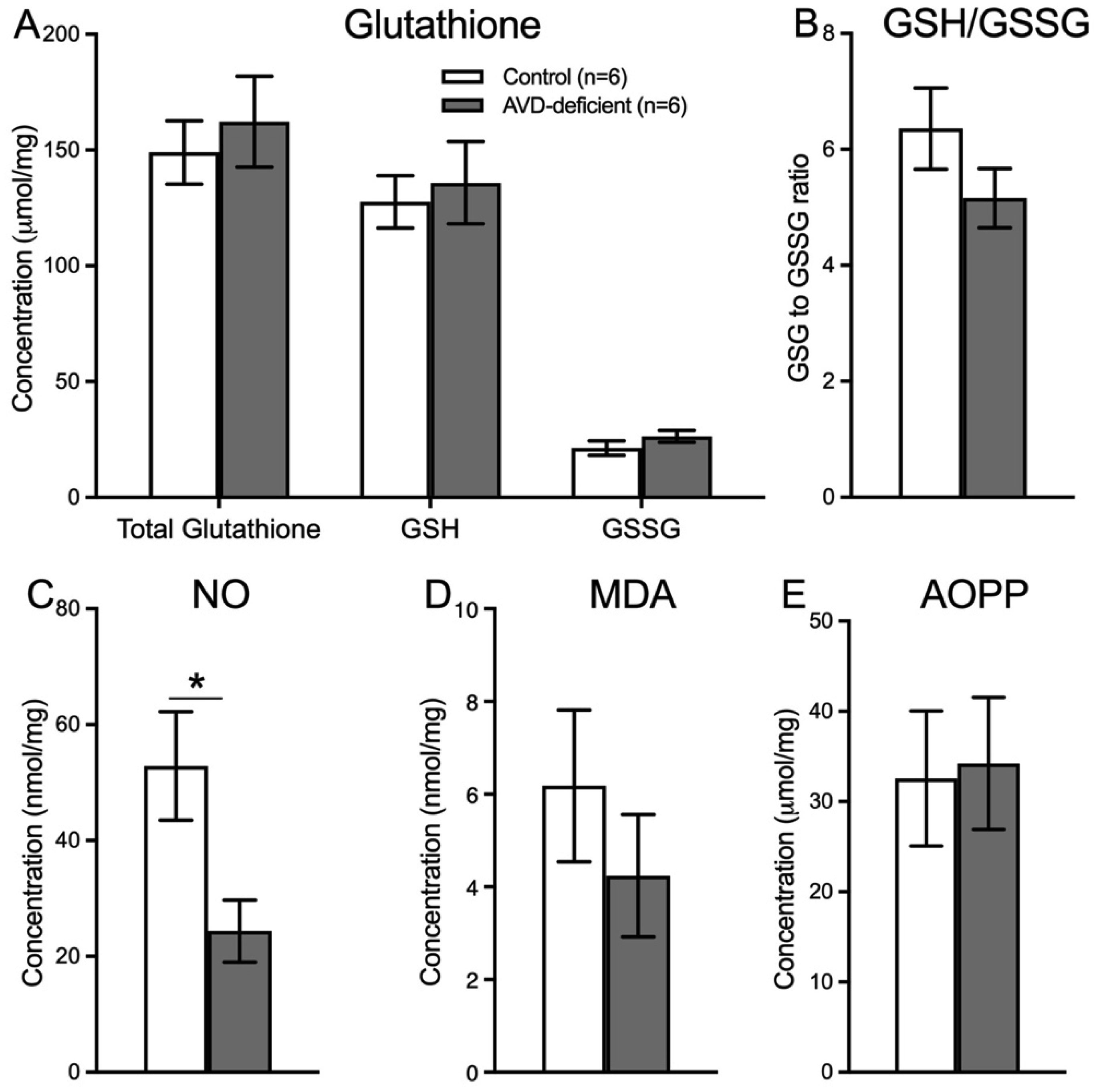
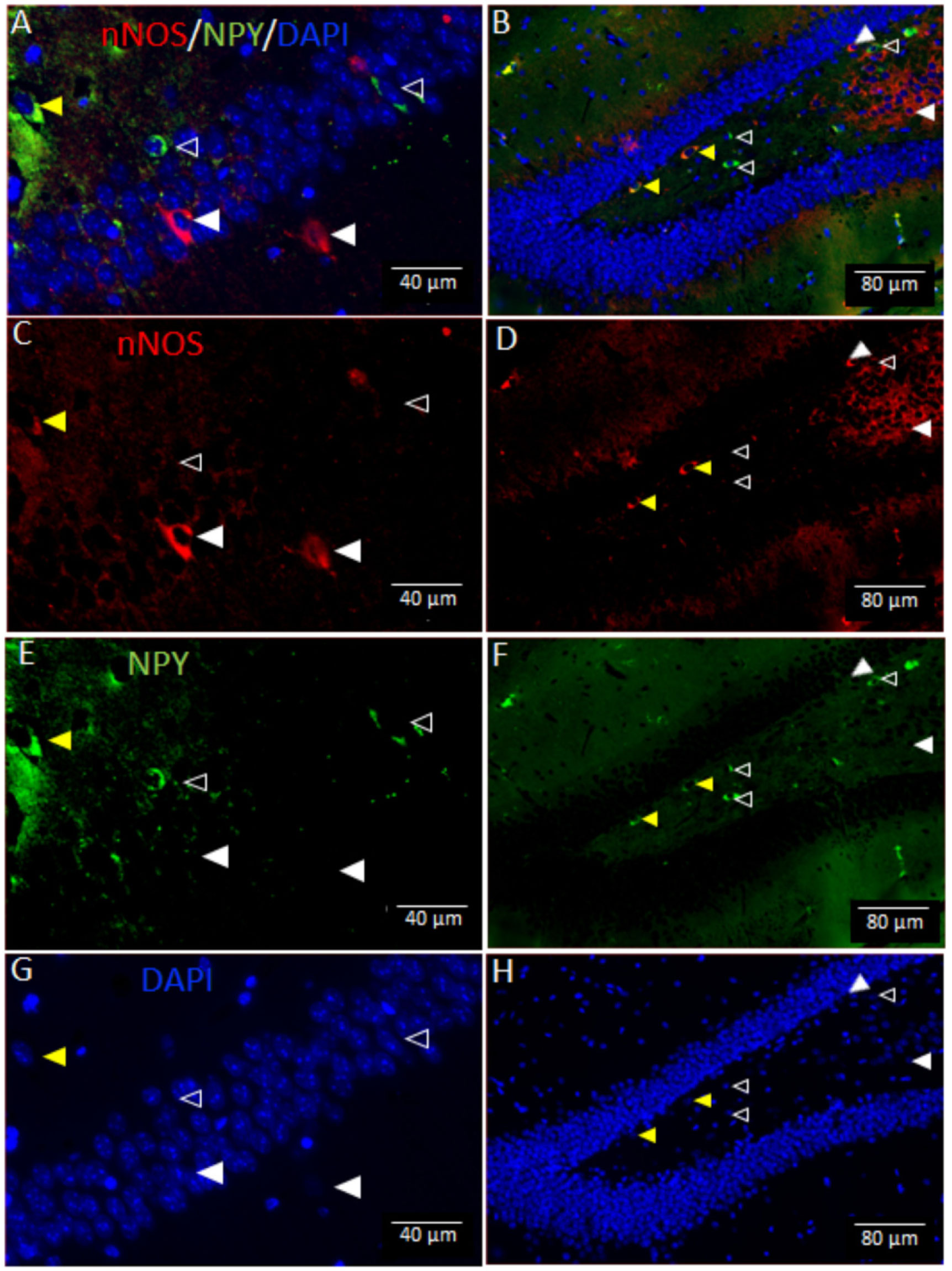
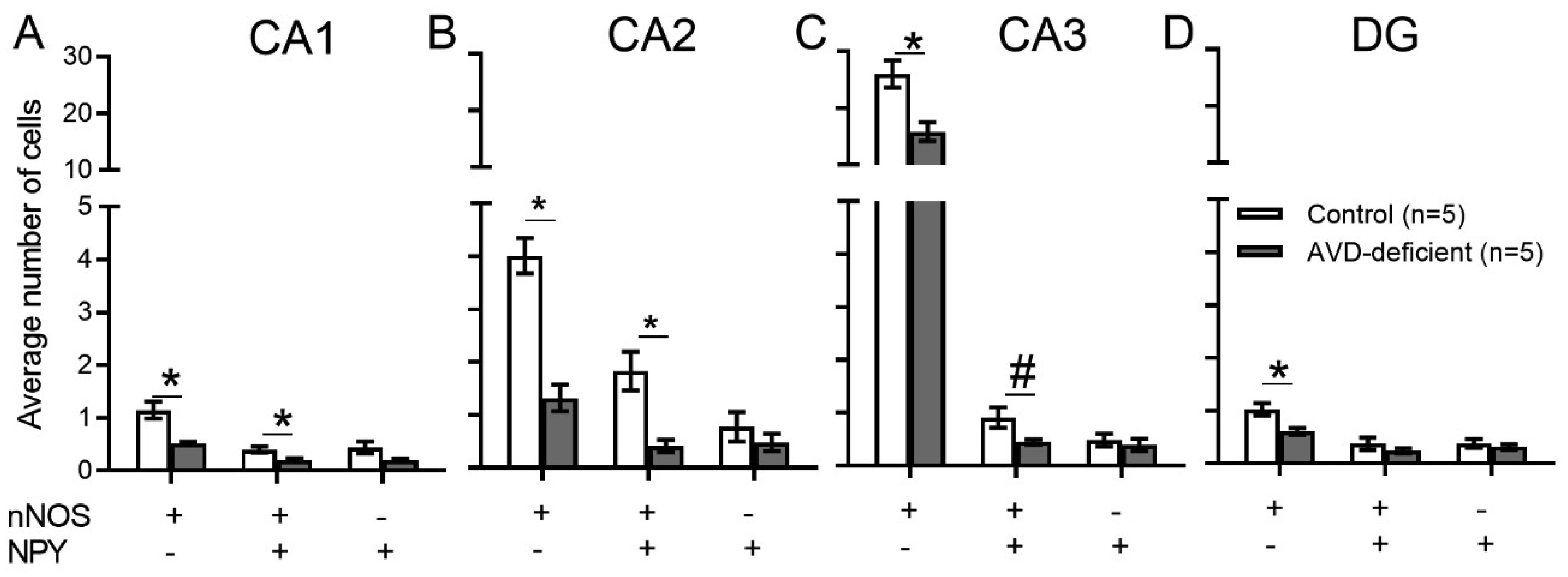
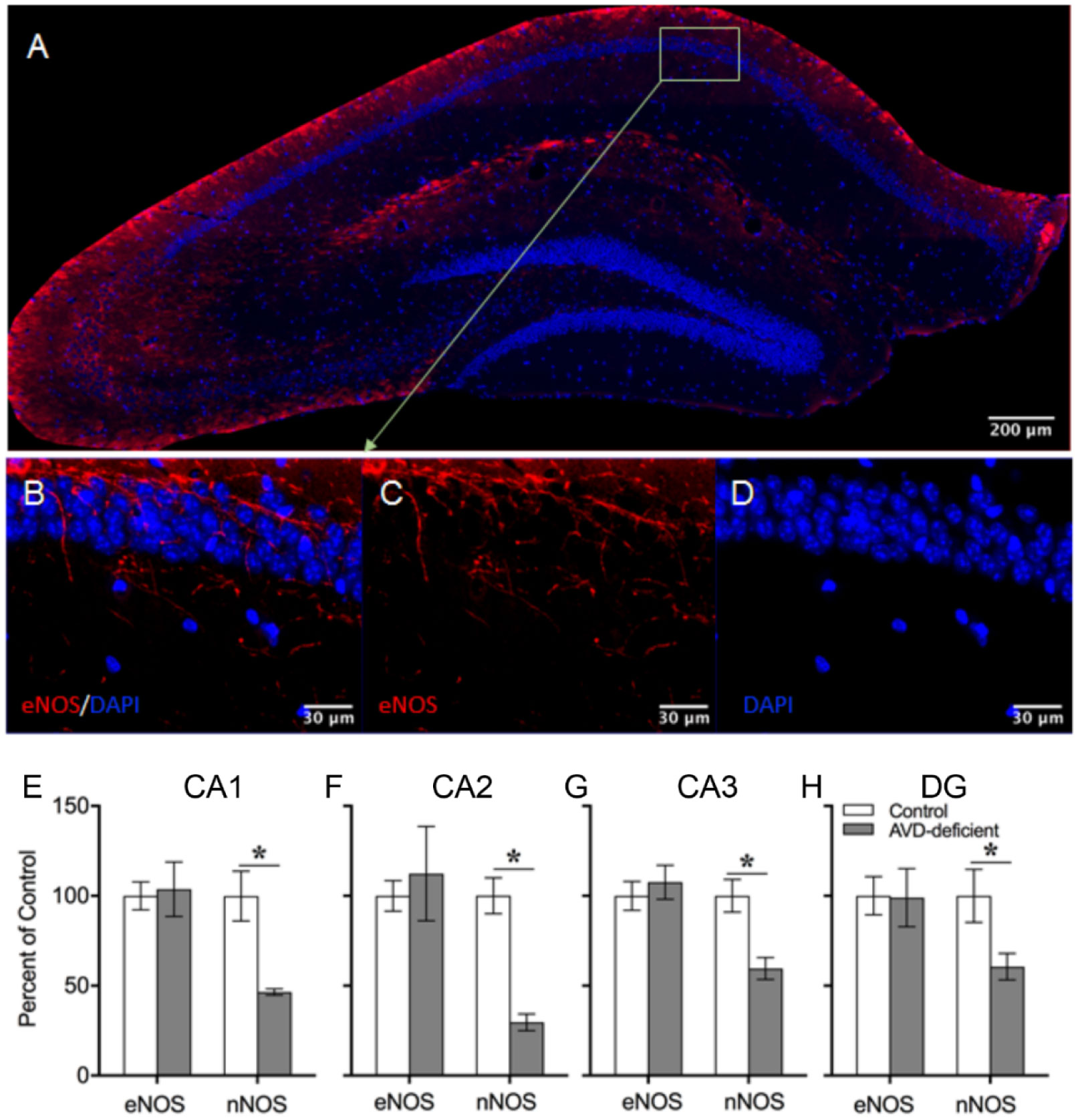
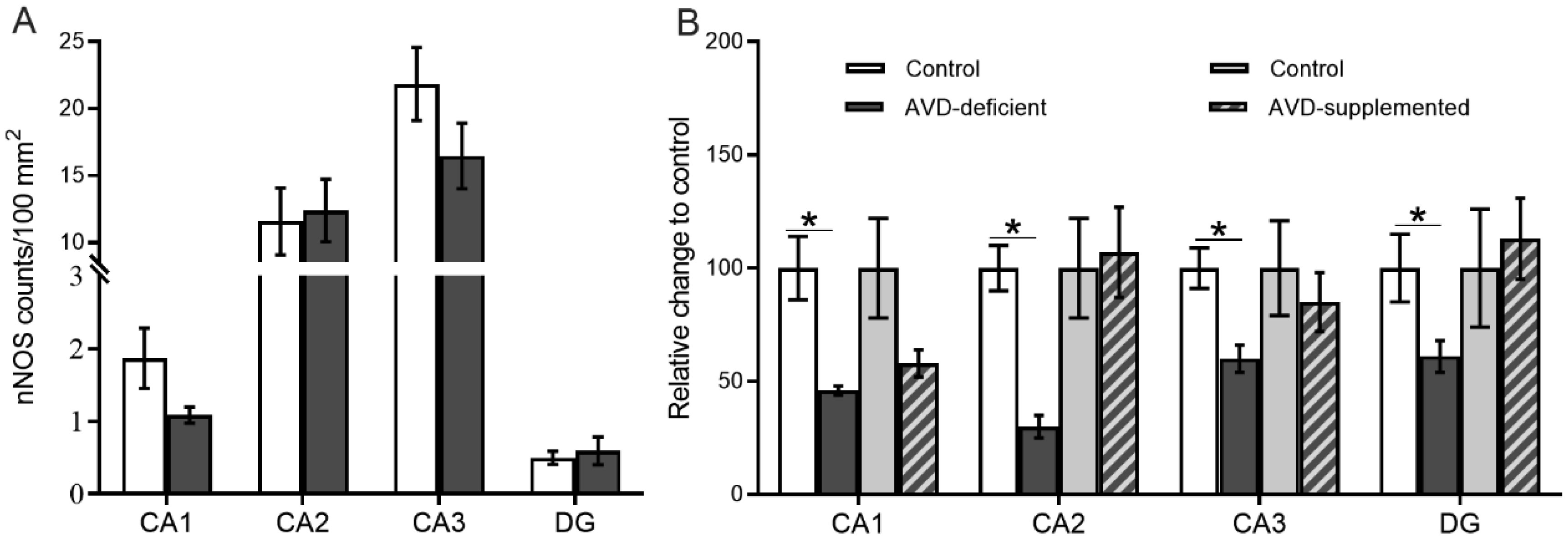
Vitamin D deficiency is prevalent in adults and is associated with cognitive impairment. However, the mechanism by which adult vitamin D (AVD) deficiency affects cognitive function remains unclear. We examined spatial memory impairment in AVD-deficient BALB/c mice and its underlying mechanism by measuring spine density, long term potentiation (LTP), nitric oxide (NO), neuronal nitric oxide synthase (nNOS), and endothelial NOS (eNOS) in the hippocampus. Adult male BALB/c mice were fed a control or vitamin D deficient diet for 20 weeks. Spatial memory performance was measured using an active place avoidance (APA) task, where AVD-deficient mice had reduced latency entering the shock zone compared to controls. We characterised hippocampal spine morphology in the CA1 and dentate gyrus (DG) and made electrophysiological recordings in the hippocampus of behaviourally naïve mice to measure LTP. We next measured NO, as well as glutathione, lipid peroxidation and oxidation of protein products and quantified hippocampal immunoreactivity for nNOS and eNOS. Spine morphology analysis revealed a significant reduction in the number of mushroom spines in the CA1 dendrites but not in the DG. There was no effect of diet on LTP. However, hippocampal NO levels were depleted whereas other oxidation markers were unaltered by AVD deficiency. We also showed a reduced nNOS, but not eNOS, immunoreactivity. Finally, vitamin D supplementation for 10 weeks to AVD-deficient mice restored nNOS immunoreactivity to that seen in in control mice. Our results suggest that lower levels of NO and reduced nNOS immunostaining contribute to hippocampal-dependent spatial learning deficits in AVD-deficient mice.
Citation: Md. Mamun Al-Amin, Robert K. P. Sullivan, Suzy Alexander, David A. Carter, DanaKai Bradford, Thomas H. J. Burne. Impaired spatial memory in adult vitamin D deficient BALB/c mice is associated with reductions in spine density, nitric oxide, and neural nitric oxide synthase in the hippocampus[J]. AIMS Neuroscience, 2022, 9(1): 31-56. doi: 10.3934/Neuroscience.2022004
Vitamin D deficiency is prevalent in adults and is associated with cognitive impairment. However, the mechanism by which adult vitamin D (AVD) deficiency affects cognitive function remains unclear. We examined spatial memory impairment in AVD-deficient BALB/c mice and its underlying mechanism by measuring spine density, long term potentiation (LTP), nitric oxide (NO), neuronal nitric oxide synthase (nNOS), and endothelial NOS (eNOS) in the hippocampus. Adult male BALB/c mice were fed a control or vitamin D deficient diet for 20 weeks. Spatial memory performance was measured using an active place avoidance (APA) task, where AVD-deficient mice had reduced latency entering the shock zone compared to controls. We characterised hippocampal spine morphology in the CA1 and dentate gyrus (DG) and made electrophysiological recordings in the hippocampus of behaviourally naïve mice to measure LTP. We next measured NO, as well as glutathione, lipid peroxidation and oxidation of protein products and quantified hippocampal immunoreactivity for nNOS and eNOS. Spine morphology analysis revealed a significant reduction in the number of mushroom spines in the CA1 dendrites but not in the DG. There was no effect of diet on LTP. However, hippocampal NO levels were depleted whereas other oxidation markers were unaltered by AVD deficiency. We also showed a reduced nNOS, but not eNOS, immunoreactivity. Finally, vitamin D supplementation for 10 weeks to AVD-deficient mice restored nNOS immunoreactivity to that seen in in control mice. Our results suggest that lower levels of NO and reduced nNOS immunostaining contribute to hippocampal-dependent spatial learning deficits in AVD-deficient mice.

| [1] |
Cieslak K, Feingold J, Antonius D, et al. (2014) Low vitamin D levels predict clinical features of schizophrenia. Schizophr Res 159: 543-545. https://doi.org/10.1016/j.schres.2014.08.031 
|
| [2] |
Chiang M, Natarajan R, Fan X (2016) Vitamin D in schizophrenia: a clinical review. Evid Based Ment Health 19: 6-9. https://doi.org/10.1136/eb-2015-102117
|
| [3] |
Fernell E, Bejerot S, Westerlund J, et al. (2015) Autism spectrum disorder and low vitamin D at birth: a sibling control study. Mol Autism 6: 3. https://doi.org/10.1186/2040-2392-6-3
|
| [4] | Saad K, Abdel-Rahman AA, Elserogy YM, et al. (2015) Vitamin D status in autism spectrum disorders and the efficacy of vitamin D supplementation in autistic children. Nutr Neurosci . https://doi.org/10.1179/1476830515Y.0000000019 |
| [5] |
Spedding S (2014) Vitamin D and depression: a systematic review and meta-analysis comparing studies with and without biological flaws. Nutrients 6: 1501-1518. https://doi.org/10.3390/nu6041501
|
| [6] | Brouwer-Brolsma EM, Dhonukshe-Rutten RA, van Wijngaarden JP, et al. (2015) Low vitamin D status is associated with more depressive symptoms in Dutch older adults. Eur J Nutr . https://doi.org/10.1007/s00394-015-0970-6 |
| [7] |
Bremner JD, Narayan M, Anderson ER, et al. (2000) Hippocampal volume reduction in major depression. Am J Psychiatry 157: 115-118. https://doi.org/10.1176/ajp.157.1.115
|
| [8] |
Li W, Li K, Guan P, et al. (2018) Volume alteration of hippocampal subfields in first-episode antipsychotic-naive schizophrenia patients before and after acute antipsychotic treatment. Neuroimage Clin 20: 169-176. https://doi.org/10.1016/j.nicl.2018.07.008
|
| [9] | Dager SR, Wang L, Friedman SD, et al. (2007) Shape mapping of the hippocampus in young children with autism spectrum disorder. AJNR Am J Neuroradiol 28: 672-677. |
| [10] |
Shivakumar V, Kalmady SV, Amaresha AC, et al. (2015) Serum vitamin D and hippocampal gray matter volume in schizophrenia. Psychiatry Res 233: 175-9. https://doi.org/10.1016/j.pscychresns.2015.06.006
|
| [11] |
Gurholt TP, Nerhus M, Osnes K, et al. (2018) Hippocampus volume reduction in psychosis spectrum could be ameliorated by vitamin D. Schizophr Res : 433-435. https://doi.org/10.1016/j.schres.2018.03.011
|
| [12] |
Al-Amin MM, Sullivan RKP, Kurniawan ND, et al. (2019) Adult vitamin D deficiency disrupts hippocampal-dependent learning and structural brain connectivity in BALB/c mice. Brain Struct Funct 224: 1315-1329. https://doi.org/10.1007/s00429-019-01840-w
|
| [13] |
Al-Amin M, Bradford D, Sullivan RKP, et al. (2019) Vitamin D deficiency is associated with reduced hippocampal volume and disrupted structural connectivity in patients with mild cognitive impairment. Hum Brain Mapp 40: 394-406. https://doi.org/10.1002/hbm.24380
|
| [14] |
Eyles DW, Smith S, Kinobe R, et al. (2005) Distribution of the vitamin D receptor and 1 alpha-hydroxylase in human brain. J Chem Neuroanat 29: 21-30. https://doi.org/10.1016/j.jchemneu.2004.08.006
|
| [15] |
Latimer CS, Brewer LD, Searcy JL, et al. (2014) Vitamin D prevents cognitive decline and enhances hippocampal synaptic function in aging rats. Proc Natl Acad Sci U S A 111: E4359-66. https://doi.org/10.1073/pnas.1404477111
|
| [16] |
Lardner AL (2015) Vitamin D and hippocampal development-the story so far. Front Mol Neurosci 8: 58. https://doi.org/10.3389/fnmol.2015.00058
|
| [17] | Al-Amin MM, Sullivan RKP, Jaeschke KN, et al. (2017) Impact of combined adult vitamin D deficiency and second-hit exposures on brain function. CAB Reviews 12: 1-16. https://doi.org/10.1079/PAVSNNR201712040 |
| [18] |
Altemus KL, Finger S, Wolf C, et al. (1987) Behavioral correlates of vitamin D deficiency. Physiol Behav 39: 435-40. https://doi.org/10.1016/0031-9384(87)90369-6
|
| [19] |
Fu L, Chen YH, Chen X, et al. (2017) Vitamin D deficiency impairs neurobehavioral development in male mice. Physiol Behav 179: 333-339. https://doi.org/10.1016/j.physbeh.2017.07.017
|
| [20] |
Vukovic J, Borlikova GG, Ruitenberg MJ, et al. (2013) Immature doublecortin-positive hippocampal neurons are important for learning but not for remembering. J Neurosci 33: 6603-13. https://doi.org/10.1523/JNEUROSCI.3064-12.2013
|
| [21] |
Willis EF, Bartlett PF, Vukovic J (2017) Protocol for Short- and Longer-term Spatial Learning and Memory in Mice. Front Behav Neurosci 11: 197. https://doi.org/10.3389/fnbeh.2017.00197
|
| [22] |
Moser MB, Trommald M, Andersen P (1994) An increase in dendritic spine density on hippocampal CA1 pyramidal cells following spatial learning in adult rats suggests the formation of new synapses. Proc Natl Acad Sci U S A 91: 12673-5. https://doi.org/10.1073/pnas.91.26.12673
|
| [23] |
Andersen P, Moser E, Moser MB, et al. (1996) Cellular correlates to spatial learning in the rat hippocampus. J Physiol Paris 90: 349. https://doi.org/10.1016/S0928-4257(97)87917-X
|
| [24] |
Mahmmoud RR, Sase S, Aher YD, et al. (2015) Spatial and Working Memory Is Linked to Spine Density and Mushroom Spines. PLoS One 10: e0139739. https://doi.org/10.1371/journal.pone.0139739
|
| [25] |
McIntire LB, Berman DE, Myaeng J, et al. (2012) Reduction of synaptojanin 1 ameliorates synaptic and behavioral impairments in a mouse model of Alzheimer's disease. J Neurosci 32: 15271-6. https://doi.org/10.1523/JNEUROSCI.2034-12.2012
|
| [26] |
Hering H, Sheng M (2001) Dendritic spines: structure, dynamics and regulation. Nat Rev Neurosci 2: 880-8. https://doi.org/10.1038/35104061
|
| [27] |
Hofer SB, Bonhoeffer T (2010) Dendritic spines: the stuff that memories are made of?. Curr Biol 20: R157-9. https://doi.org/10.1016/j.cub.2009.12.040
|
| [28] |
Salami M, Talaei SA, Davari S, et al. (2012) Hippocampal long term potentiation in rats under different regimens of vitamin D: an in vivo study. Neurosci Let 509: 56-9. https://doi.org/10.1016/j.neulet.2011.12.050
|
| [29] |
Bashir ZI, Bortolotto ZA, Davies CH, et al. (1993) Induction of LTP in the hippocampus needs synaptic activation of glutamate metabotropic receptors. Nature 363: 347-50. https://doi.org/10.1038/363347a0
|
| [30] |
Groves NJ, Bradford D, Sullivan RK, et al. (2016) Behavioural Effects of Adult Vitamin D Deficiency in BALB/c Mice Are not Associated with Proliferation or Survival of Neurons in the Adult Hippocampus. PLoS One 11: e0152328. https://doi.org/10.1371/journal.pone.0152328
|
| [31] |
Groves NJ, Kesby JP, Eyles DW, et al. (2013) Adult vitamin D deficiency leads to behavioural and brain neurochemical alterations in C57BL/6J and BALB/c mice. Behav Brain Res 241: 120-31. https://doi.org/10.1016/j.bbr.2012.12.001
|
| [32] |
Bao BY, Ting HJ, Hsu JW, et al. (2008) Protective role of 1 alpha, 25-dihydroxyvitamin D3 against oxidative stress in nonmalignant human prostate epithelial cells. Int J Cancer 122: 2699-706. https://doi.org/10.1002/ijc.23460
|
| [33] |
Garcion E, Sindji L, Leblondel G, et al. (1999) 1,25-dihydroxyvitamin D3 regulates the synthesis of gamma-glutamyl transpeptidase and glutathione levels in rat primary astrocytes. J Neurochem 73: 859-66. https://doi.org/10.1046/j.1471-4159.1999.0730859.x
|
| [34] |
Bredt DS, Glatt CE, Hwang PM, et al. (1991) Nitric oxide synthase protein and mRNA are discretely localized in neuronal populations of the mammalian CNS together with NADPH diaphorase. Neuron 7: 615-24. https://doi.org/10.1016/0896-6273(91)90374-9
|
| [35] |
Kuriyama K, Ohkuma S (1995) Role of nitric oxide in central synaptic transmission: effects on neurotransmitter release. Jpn J Pharmacol 69: 1-8. https://doi.org/10.1254/jjp.69.1
|
| [36] |
Bradley SA, Steinert JR (2016) Nitric Oxide-Mediated Posttranslational Modifications: Impacts at the Synapse. Oxid Med Cell Longev 2016: 5681036. https://doi.org/10.1155/2016/5681036
|
| [37] |
Hardingham N, Dachtler J, Fox K (2013) The role of nitric oxide in pre-synaptic plasticity and homeostasis. Front Cell Neurosci 7: 190. https://doi.org/10.3389/fncel.2013.00190
|
| [38] |
Lange MD, Doengi M, Lesting J, et al. (2012) Heterosynaptic long-term potentiation at interneuron-principal neuron synapses in the amygdala requires nitric oxide signalling. J Physiol 590: 131-43. https://doi.org/10.1113/jphysiol.2011.221317
|
| [39] |
O'Dell TJ, Hawkins RD, Kandel ER, et al. (1991) Tests of the roles of two diffusible substances in long-term potentiation: evidence for nitric oxide as a possible early retrograde messenger. Proc Natl Acad Sci U S A 88: 11285-9. https://doi.org/10.1073/pnas.88.24.11285
|
| [40] |
Andrukhova O, Slavic S, Zeitz U, et al. (2014) Vitamin D is a regulator of endothelial nitric oxide synthase and arterial stiffness in mice. Mol Endocrinol 28: 53-64. https://doi.org/10.1210/me.2013-1252
|
| [41] |
Rockett KA, Brookes R, Udalova I, et al. (1998) 1,25-Dihydroxyvitamin D3 induces nitric oxide synthase and suppresses growth of Mycobacterium tuberculosis in a human macrophage-like cell line. Infect Immun 66: 5314-21. https://doi.org/10.1128/IAI.66.11.5314-5321.1998
|
| [42] |
Gotzsche CR, Woldbye DP (2016) The role of NPY in learning and memory. Neuropeptides 55: 79-89. https://doi.org/10.1016/j.npep.2015.09.010
|
| [43] |
dos Santos VV, Santos DB, Lach G, et al. (2013) Neuropeptide Y (NPY) prevents depressive-like behavior, spatial memory deficits and oxidative stress following amyloid-beta (Abeta(1-40)) administration in mice. Behav Brain Res 244: 107-15. https://doi.org/10.1016/j.bbr.2013.01.039
|
| [44] |
Tricoire L., Pelkey K.A., Daw M.I., et al. (2010) Common origins of hippocampal Ivy and nitric oxide synthase expressing neurogliaform cells. J Neurosci 30: 2165-76. https://doi.org/10.1523/JNEUROSCI.5123-09.2010
|
| [45] |
Blackshaw S, Eliasson MJ, Sawa A, et al. (2003) Species, strain and developmental variations in hippocampal neuronal and endothelial nitric oxide synthase clarify discrepancies in nitric oxide-dependent synaptic plasticity. Neuroscience 119: 979-90. https://doi.org/10.1016/S0306-4522(03)00217-3
|
| [46] |
Aoki C, Rhee J, Lubin M, et al. (1997) NMDA-R1 subunit of the cerebral cortex co-localizes with neuronal nitric oxide synthase at pre- and postsynaptic sites and in spines. Brain Res 750: 25-40. https://doi.org/10.1016/S0006-8993(96)01147-X
|
| [47] |
Lobellova V, Brichtova E, Petrasek T, et al. (2015) Higher doses of (+)MK-801 (dizocilpine) induced mortality and procedural but not cognitive deficits in delayed testing in the active place avoidance with reversal on the Carousel. Physiol Res 64: 269-275. https://doi.org/10.33549/physiolres.932832
|
| [48] |
Stuchlik A, Petrasek T, Prokopova I, et al. (2013) Place avoidance tasks as tools in the behavioral neuroscience of learning and memory. Physiol Res 62 Suppl 1: S1-s19. https://doi.org/10.33549/physiolres.932635
|
| [49] |
Wesierska M, Dockery C, Fenton AA (2005) Beyond memory, navigation, and inhibition: behavioral evidence for hippocampus-dependent cognitive coordination in the rat. J Neurosci 25: 2413-9. https://doi.org/10.1523/JNEUROSCI.3962-04.2005
|
| [50] |
Das G, Reuhl K, Zhou R (2013) The Golgi-Cox method. Methods Mol Biol 1018: 313-21. https://doi.org/10.1007/978-1-62703-444-9_29
|
| [51] |
Carpenter AE, Jones TR, Lamprecht MR, et al. (2006) CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biol 7: R100. https://doi.org/10.1186/gb-2006-7-10-r100
|
| [52] |
Dickstein DL, Dickstein DR, Janssen WG, et al. (2016) Automatic Dendritic Spine Quantification from Confocal Data with Neurolucida 360. Curr Protoc Neurosci 77: 1.27.1-1.27.21. https://doi.org/10.1002/cpns.16
|
| [53] | Tracey WR, Tse J, Carter G (1995) Lipopolysaccharide-induced changes in plasma nitrite and nitrate concentrations in rats and mice: pharmacological evaluation of nitric oxide synthase inhibitors. J Pharmacol Exp Ther 272: 1011-5. |
| [54] |
Al-Amin MM, Alam T, Hasan SM, et al. (2016) Prenatal maternal lipopolysaccharide administration leads to age- and region-specific oxidative stress in the early developmental stage in offspring. Neuroscience 318: 84-93. https://doi.org/10.1016/j.neuroscience.2016.01.002
|
| [55] |
Sun J, Zhang X, Broderick M, et al. (2003) Measurement of nitric oxide production in biological systems by using Griess reaction assay. Sensors 3: 276-284. https://doi.org/10.3390/s30800276
|
| [56] |
Rahman I, Kode A, Biswas SK (2006) Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat Protoc 1: 3159-65. https://doi.org/10.1038/nprot.2006.378
|
| [57] |
Witko-Sarsat V, Friedlander M, Capeillere-Blandin C, et al. (1996) Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int 49: 1304-13. https://doi.org/10.1038/ki.1996.186
|
| [58] |
Al-Amin MM, Hasan SM, Alam T, et al. (2014) Tadalafil enhances working memory, and reduces hippocampal oxidative stress in both young and aged mice. Eur J Pharmacol 745: 84-90. https://doi.org/10.1016/j.ejphar.2014.10.026
|
| [59] |
Niehaus WG, Samuelsson B (1968) Formation of malonaldehyde from phospholipid arachidonate during microsomal lipid peroxidation. Eur J Biochem 6: 126-130. https://doi.org/10.1111/j.1432-1033.1968.tb00428.x
|
| [60] |
Al-Amin MM, Sultana R, Sultana S, et al. (2016) Astaxanthin ameliorates prenatal LPS-exposed behavioral deficits and oxidative stress in adult offspring. BMC Neurosci 17: 11. https://doi.org/10.1186/s12868-016-0245-z
|
| [61] |
Eyles D, Anderson C, Ko P, et al. (2009) A sensitive LC/MS/MS assay of 25OH vitamin D3 and 25OH vitamin D2 in dried blood spots. Clin Chim Acta 403: 145-151. https://doi.org/10.1016/j.cca.2009.02.005
|
| [62] |
Leuner B, Falduto J, Shors TJ (2003) Associative memory formation increases the observation of dendritic spines in the hippocampus. J Neurosci 23: 659-665. https://doi.org/10.1523/JNEUROSCI.23-02-00659.2003
|
| [63] |
Diamond DM, Campbell AM, Park CR, et al. (2006) Influence of predator stress on the consolidation versus retrieval of long-term spatial memory and hippocampal spinogenesis. Hippocampus 16: 571-6. https://doi.org/10.1002/hipo.20188
|
| [64] |
Tronel S, Fabre A, Charrier V, et al. (2010) Spatial learning sculpts the dendritic arbor of adult-born hippocampal neurons. Proc Natl Acad Sci U S A 107: 7963-8. https://doi.org/10.1073/pnas.0914613107
|
| [65] |
Beltran-Campos V, Prado-Alcala RA, Leon-Jacinto U, et al. (2011) Increase of mushroom spine density in CA1 apical dendrites produced by water maze training is prevented by ovariectomy. Brain Res 1369: 119-30. https://doi.org/10.1016/j.brainres.2010.10.105
|
| [66] |
O'Malley A, O'Connell C, Murphy KJ, et al. (2000) Transient spine density increases in the mid-molecular layer of hippocampal dentate gyrus accompany consolidation of a spatial learning task in the rodent. Neuroscience 99: 229-32. https://doi.org/10.1016/S0306-4522(00)00182-2
|
| [67] |
Eyre MD, Richter-Levin G, Avital A, et al. (2003) Morphological changes in hippocampal dentate gyrus synapses following spatial learning in rats are transient. Eur J Neurosci 17: 1973-80. https://doi.org/10.1046/j.1460-9568.2003.02624.x
|
| [68] |
Pavlowsky A, Wallace E, Fenton AA, et al. (2017) Persistent modifications of hippocampal synaptic function during remote spatial memory. Neurobiol Learn Mem 138: 182-197. https://doi.org/10.1016/j.nlm.2016.08.015
|
| [69] |
Graham BP, Saudargiene A, Cobb S (2014) Spine head calcium as a measure of summed postsynaptic activity for driving synaptic plasticity. Neural Comput 26: 2194-222. https://doi.org/10.1162/NECO_a_00640
|
| [70] |
Harris KM (1999) Calcium from internal stores modifies dendritic spine shape. Proc Natl Acad Sci U S A 96: 12213-5. https://doi.org/10.1073/pnas.96.22.12213
|
| [71] |
Citri A, Malenka RC (2008) Synaptic plasticity: multiple forms, functions, and mechanisms. Neuropsychopharmacology 33: 18-41. https://doi.org/10.1038/sj.npp.1301559
|
| [72] |
Collin C, Miyaguchi K, Segal M (1997) Dendritic spine density and LTP induction in cultured hippocampal slices. J Neurophysiol 77: 1614-1623. https://doi.org/10.1152/jn.1997.77.3.1614
|
| [73] |
Steele P, Mauk MD (1999) Inhibitory control of LTP and LTD: stability of synapse strength. J Neurophysiol 81: 1559-1566. https://doi.org/10.1152/jn.1999.81.4.1559
|
| [74] |
Huang TT, Leu D, Zou Y (2015) Oxidative stress and redox regulation on hippocampal-dependent cognitive functions. Arch Biochem Biophys 576: 2-7. https://doi.org/10.1016/j.abb.2015.03.014
|
| [75] |
Chang JM, Kuo MC, Kuo HT, et al. (2004) 1-alpha,25-Dihydroxyvitamin D3 regulates inducible nitric oxide synthase messenger RNA expression and nitric oxide release in macrophage-like RAW 264.7 cells. J Lab Clin Med 143: 14-22. https://doi.org/10.1016/j.lab.2003.08.002
|
| [76] |
Zou LB, Yamada K, Tanaka T, et al. (1998) Nitric oxide synthase inhibitors impair reference memory formation in a radial arm maze task in rats. Neuropharmacology 37: 323-330. https://doi.org/10.1016/S0028-3908(98)00042-2
|
| [77] |
Moosavi M, Abbasi L, Zarifkar A, et al. (2014) The role of nitric oxide in spatial memory stages, hippocampal ERK and CaMKII phosphorylation. Pharmacol Biochem Behav 122: 164-172. https://doi.org/10.1016/j.pbb.2014.03.021
|
| [78] |
Harooni HE, Naghdi N, Sepehri H, et al. (2009) The role of hippocampal nitric oxide (NO) on learning and immediate, short- and long-term memory retrieval in inhibitory avoidance task in male adult rats. Behav Brain Res 201: 166-172. https://doi.org/10.1016/j.bbr.2009.02.011
|
| [79] |
Bon CL, Garthwaite J (2003) On the role of nitric oxide in hippocampal long-term potentiation. J Neurosci 23: 1941-1948. https://doi.org/10.1523/JNEUROSCI.23-05-01941.2003
|
| [80] |
Sase A, Nawaratna G, Hu S, et al. (2016) Decreased hippocampal homoarginine and increased nitric oxide and nitric oxide synthase levels in rats parallel training in a radial arm maze. Amino Acids 48: 2197-2204. https://doi.org/10.1007/s00726-016-2251-y
|
| [81] |
Wang S, Pan DX, Wang D, et al. (2014) Nitric oxide facilitates active avoidance learning via enhancement of glutamate levels in the hippocampal dentate gyrus. Behav Brain Res 271: 177-183. https://doi.org/10.1016/j.bbr.2014.06.011
|
| [82] |
Son H, Hawkins RD, Martin K, et al. (1996) Long-term potentiation is reduced in mice that are doubly mutant in endothelial and neuronal nitric oxide synthase. Cell 87: 1015-1023. https://doi.org/10.1016/S0092-8674(00)81796-1
|
| [83] |
Calabrese V, Mancuso C, Calvani M, et al. (2007) Nitric oxide in the central nervous system: neuroprotection versus neurotoxicity. Nat Rev Neurosci 8: 766-775. https://doi.org/10.1038/nrn2214
|
| [84] |
Bredt DS (1999) Endogenous nitric oxide synthesis: biological functions and pathophysiology. Free Radic Res 31: 577-596. https://doi.org/10.1080/10715769900301161
|
| [85] |
Koprowski H, Zheng YM, Heber-Katz E, et al. (1993) In vivo expression of inducible nitric oxide synthase in experimentally induced neurologic diseases. Proc Natl Acad Sci U S A 90: 3024-3027. https://doi.org/10.1073/pnas.90.7.3024
|
| [86] |
Tricoire L, Vitalis T (2012) Neuronal nitric oxide synthase expressing neurons: a journey from birth to neuronal circuits. Front Neural Circuits 6: 82. https://doi.org/10.3389/fncir.2012.00082
|
| [87] |
Zhang S, Chen J, Wang S (1998) Spatial learning and memory induce up-regulation of nitric oxide-producing neurons in rat brain. Brain Res 801: 101-106. https://doi.org/10.1016/S0006-8993(98)00564-2
|
| [88] |
Brenman JE, Chao DS, Gee SH, et al. (1996) Interaction of nitric oxide synthase with the postsynaptic density protein PSD-95 and alpha1-syntrophin mediated by PDZ domains. Cell 84: 757-767. https://doi.org/10.1016/S0092-8674(00)81053-3
|
| [89] |
Burette A, Zabel U, Weinberg RJ, et al. (2002) Synaptic localization of nitric oxide synthase and soluble guanylyl cyclase in the hippocampus. J Neurosci 22: 8961-8970. https://doi.org/10.1523/JNEUROSCI.22-20-08961.2002
|
| [90] |
Doyle CA, Slater P (1997) Localization of neuronal and endothelial nitric oxide synthase isoforms in human hippocampus. Neuroscience 76: 387-395. https://doi.org/10.1016/S0306-4522(96)00297-7
|
| [91] |
Caviedes A, Varas-Godoy M, Lafourcade C, et al. (2017) Endothelial Nitric Oxide Synthase Is Present in Dendritic Spines of Neurons in Primary Cultures. Front Cell Neurosci 11: 180. https://doi.org/10.3389/fncel.2017.00180
|
| [92] |
Fuentealba P, Begum R, Capogna M, et al. (2008) Ivy cells: a population of nitric-oxide-producing, slow-spiking GABAergic neurons and their involvement in hippocampal network activity. Neuron 57: 917-929. https://doi.org/10.1016/j.neuron.2008.01.034
|
| [93] |
Somogyi J, Szabo A, Somogyi P, et al. (2012) Molecular analysis of ivy cells of the hippocampal CA1 stratum radiatum using spectral identification of immunofluorophores. Front Neural Circuits 6: 35. https://doi.org/10.3389/fncir.2012.00035
|
| [94] |
Morley JE, Alshaher MM, Farr SA, et al. (1999) Leptin and neuropeptide Y (NPY) modulate nitric oxide synthase: further evidence for a role of nitric oxide in feeding. Peptides 20: 595-600. https://doi.org/10.1016/S0196-9781(99)00012-1
|
| [95] |
Jedlicka P, Vlachos A, Schwarzacher SW, et al. (2008) A role for the spine apparatus in LTP and spatial learning. Behav Brain Res 192: 12-19. https://doi.org/10.1016/j.bbr.2008.02.033
|
| [96] |
Toni N, Buchs PA, Nikonenko I, et al. (1999) LTP promotes formation of multiple spine synapses between a single axon terminal and a dendrite. Nature 402: 421-425. https://doi.org/10.1038/46574
|
| [97] |
Malen PL, Chapman PF (1997) Nitric oxide facilitates long-term potentiation, but not long-term depression. J Neurosci 17: 2645-2651. https://doi.org/10.1523/JNEUROSCI.17-07-02645.1997
|
| [98] |
Bohme GA, Bon C, Stutzmann JM, et al. (1991) Possible involvement of nitric oxide in long-term potentiation. Eur J Pharmacol 199: 379-381. https://doi.org/10.1016/0014-2999(91)90505-K
|
| [99] |
Phillips KG, Hardingham NR, Fox K (2008) Postsynaptic action potentials are required for nitric-oxide-dependent long-term potentiation in CA1 neurons of adult GluR1 knock-out and wild-type mice. J Neurosci 28: 14031-14041. https://doi.org/10.1523/JNEUROSCI.3984-08.2008
|
| [100] |
Wilson RI, Yanovsky J, Godecke A, et al. (1997) Endothelial nitric oxide synthase and LTP. Nature 386: 338. https://doi.org/10.1038/386338a0
|
 neurosci-09-01-004-s001.pdf neurosci-09-01-004-s001.pdf |
 |
Figures(9) / Tables(1)

Md. Mamun Al-Amin, Robert K. P. Sullivan, Suzy Alexander, David A. Carter, DanaKai Bradford, Thomas H. J. Burne. Impaired spatial memory in adult vitamin D deficient BALB/c mice is associated with reductions in spine density, nitric oxide, and neural nitric oxide synthase in the hippocampus[J]. AIMS Neuroscience, 2022, 9(1): 31-56. doi: 10.3934/Neuroscience.2022004







 DownLoad:
DownLoad: